


| name | Amanita alliacea |
| name status | nomen acceptum |
| author | (Murrill) Murrill |
| english name | "Garlic-Odored Limbed-Lepidella" |
| intro | The following is taken from Bas' 1969 thesis on Amanita section Lepidella. |
| cap |
The cap of Amanita alliacea is about 80 mm wide, convex, white, dry, shiny, with an appendiculate, nonsulcate margin. The cap is decorated with scattered, vague, thin, felted-subpulverulent, white patches of volva. |
| gills |
The gills are rather crowded, adnate to nearly free, narrow, and white. The short gills are truncate to obliquely truncate to attenuate. |
| stem |
The stem is 100 × 20 mm, tapering upward, solid, white, floccose, subannulate, with a volval limb collapsed against the base of the stem. |
| spores |
The spores measure (12-) 13 - 14.5 × 4 - 4.5 µm and are amyloid and bacilliform. Clamps are absent at bases of basidia. |
| discussion |
The type of A. alliacea was collected under oak; other collections were made in "mixed woods." This species was described from Gainesville, Florida, U.S.A. Bas (1969) says that the two Murrill collections (including the type) that he examined did not conform completely with the original description. Moreover, Bas states that his study showed the odor of garlic was the only character separating the material he revised from A. roanokensis Coker. However, Murrill had reported A. alliacea as very common in the Gainesville area. Bas, therefore, felt that he should leave the species standing for the time being since Murrill apparently had familiarity with it, but had never suggested that it was the same as A. watsoniana (Murrill's name for A. roanokensis).—R. E. Tulloss |
| brief editors | RET |
| name | Amanita alliacea | ||||||||
| author | (Murrill) Murrill. 1941. Mycologia 33: 448. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Garlic-Odored Limbed-Lepidella" | ||||||||
| synonyms |
≡Venenarius alliaceus Murrill. 1941. Mycologia 33: 434. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 284046, 291937 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | FLAS | ||||||||
| type studies | Jenkins. 1979. Mycotaxon 10: 176. | ||||||||
| revisions | Bas. 1969. Persoonia 5: 552, figs. 369-373. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following text is derived primarily from the revision of Bas (1969), with other sources as noted. Bas (1969): Basidiomes medium-sized, rather slender. | ||||||||
| pileus | Bas (1969): ca. 80 mm wide, white, dry (probably subviscid when moist), shiny; context white, unchanging; margin appendiculate, nonstriate; universal veil as vague thin subpulverulent white patches 4 - 12 mm wide. | ||||||||
| lamellae | Bas (1969): adnate to narrowly free, rather crowded, white, narrow, with entire edge; lamellulae with shorter ones truncate to obliquely truncate, with longer attenuate. | ||||||||
| stipe | Bas (1969): ca. 100 × 20 mm, white, narrowing upward,floccose; bulb broadly fusiform, slightly rooting, 40 × 35 mm; context solid, white, unchanging; partial veil submembranous, fragile, well-formed at first, soon tearing, leaving annular zone or disappearing; universal veil as limbate volva, with limb up to 20 mm high, collapsing on stipe base according to Murrill, in exsicata seen only in young specimen (as some vague felted subpulverulent patches at top of bulb and base of stipe. | ||||||||
| odor/taste | Bas (1969): Odor strongly of garlic. Taste not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | Bas (1969): . | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | Bas (1969): bilateral; "probably without terminal inflated cells." | ||||||||
| subhymenium | Bas (1969): "ramose to subcellular." | ||||||||
| basidia | Bas (1969): 40 - 55 × 9 - 11 μm, 4-sterigmate, with many having refractive contents; "clampless." | ||||||||
| universal veil | Bas (1969): On pileus: "yellowish in alkaline solution"; filamentous hyphae 2 - 6 μm wide, abundant, sometimes with rather thickened walls, dominant on patch surfaces; inflated cells globose to ovoid to broadly ellipsoid, 25 - 60 × 20 - 50 μm, terminal singly (mostly) or in short rows, infrequent near patch surfaces. On stipe base: "in young specimen similar to those on cap." | ||||||||
| stipe context | Bas (1969): longitudinally acrophysalidic. | ||||||||
| columnella context | double click in markup mode to edit. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | Bas (1969): "difficult to study" [due to condition], "probably consisting mainly of broadly clavate to subglobose cells about 25 - 40 × 12 - 25 μm." | ||||||||
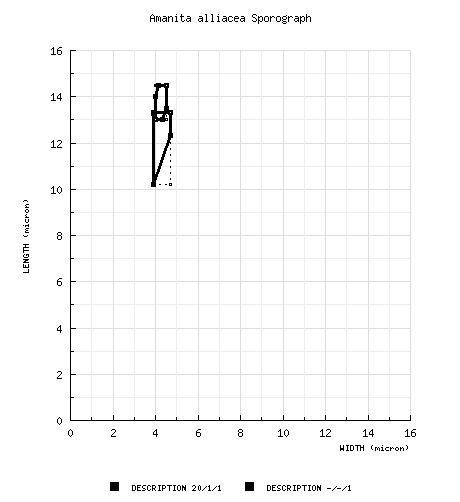
| basidiospores |
Bas (1969): [20/1/1] (12.0-) 13.0 - 14.5 × 4.0 - 4.5 μm, (Q = (1.70-) 3.0 - 3.50; Q = 3.20), slightly yellowish, thin-walled, amyloid, bacilliform, rarely cylindric or elongate, sometimes with very minute amyloid warts; apiculus not described; contents subgranular-refractive; color in deposit not recorded. from type study of Jenkins (1979): [-/-/1] 10.2 - 13.3 × (3.1-) 3.9 - 4.7 μm, (Q = 2.62 - 3.4; Q' = 2.97), hyaline, thin-walled, amyloid, cylindric to bacilliform, often adaxially flattened; apiculus sublateral, cylindric; contents guttulate; color in deposit not recorded. | ||||||||
| ecology | Bas (1969): Terrestrial. Under Quercus (type) or in mixed woods (Murrill 1948: 107). | ||||||||
| material examined |
Bas (1969): U.S.A.:
FLORIDA—Alachua Co. - Tung-oil Mill ca. Gainesville, 18.vi.1938 W. A. Murrill F 16418 (holotype, FLAS). from type study of Jenkins (1979): U. S. A.: FLORIDA— Alachua Co. - Gainesville, 18.vi.1938 W. A. Murrill F 16418 (holotype, FLAS). | ||||||||
| discussion |
Bas (1969): "In Murrill's field notes the sketch shows a fungus with a rather high, collar-like volval limb on the bulb; in his description the limb of the volva is described as being collapsed against the base of the stem. in the type material is a volval limb completely lacking while only in the youngest specimen are vague patches of volva found appressed against the base of the stem. This is all somewhat confusing. A study of the microscopic structure of the patches of volva on the cap, however, revealed that it is similar to that of A. roanokensis. Presumably in the present species a very fragile limb is present, just as in A. roanokensis, but it may fall away easily, which has in fact happened in the type specimens. "Amanita alliacea, with its very long, slender spores and submembranous volva, is very close, perhaps too close, to A. roanokensis, which occurs in the same area. Actually the strong garlicky smell, very uncommon in the genus Amanita, seems to be the only character separating the two, but I studied only the two type specimens of A. alliacea and these do not fully agree with the protologue (see observations above). Moreover, according to Murrill (1948: 107; 1951: 20) the species occurs frequently in Florida, which means that Murrill knew it rather well. Yet he did not suggest a close relationship with A. roanokensis (his A. watsonianus). Therefore I think that any proposal to change the taxonomic position of A. alliacea should await further information." | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita alliacea |
| bottom links |
[ Keys & Checklists ] [ E. Texas & Gulf Coast list ] |
| name | Amanita alliacea |
| bottom links |
[ Keys & Checklists ] [ E. Texas & Gulf Coast list ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.