


| name | Amanita roanokensis |
| name status | nomen acceptum |
| author | Coker |
| english name | "Roanoke Limbed Lepidella" |
| images |
 1. Amanita roanokensis, eastern Texas, U.S.A.  2. Amanita roanokensis, eastern Texas, U.S.A. |
| intro |
Amanita roanokensis has very narrow spores, which commonly may be more than 4 times as long as they are wide. |
| cap |
The cap is starkly white and 50 - 120 mm wide, convex to plano-convex, sometimes with a slight umbo, white, cream in the center, probably subviscid, with a nonsulcate, appendiculate margin. The cap is scattered with rather small to large, thin, flat, white, submembranous patches of volva. |
| gills |
The gills are white, adnate or adnexed, and moderately crowded to crowded to rather distant, narrow to moderately broad, and white to pallid. The short gills are obliquely truncate to attenuate. |
| stem |
The usually exannulate stipe is 70 - 140 × 7 - 20 mm and, subcylindrical or attenuate upward, solid, white, and (at first) flocculose; at the stipe base is a spindle-shaped, somewhat rooting bulb. A weak annulus may be noticed in young fruiting bodies. There is a distinct limbate volva at the top of the stipe's basal bulb, but this may be hard to see when it collapses on the stipe. |
| spores |
The spores measure (12.0-) 12.8 - 17.1 (-19.5) × 3.6 - 4.9 (-5.0) µm and are amyloid and cylindrical to bacilliform. Clamps are not found at bases of basidia. |
| discussion |
Amanita roanokensis has an odor that is pleasant or "like cooking meat" at first, but later it is described as "like carrion" or "old bones." This species is often associated with oak and pine. It is moderately common in the southern part of the sandy Atlantic coastal plain of the U.S. and along the U.S. Gulf Coast. Bas placed the present species in his stirps Roanokensis along with A. inodora (Murrill) Bas and A. alliacea (Murrill) Murrill. For a brief statement concerning the related stirpes of Amanita subsection Limbatulae, see A. limbatula Bas.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita roanokensis | ||||||||
| author | Coker. 1927. J. Elisha Mitchell Scient. Soc. 47: 141, pl. 2 (fig. 6). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Roanoke Limbed Lepidella" | ||||||||
| synonyms |
≡Venenarius roanokensis (Coker) Murrill. 1948. Lloydia 11: 103.
≡Amidella peckiana f. roanokensis (Coker) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 77.
=Venenarius watsonianus Murrill. 1945c ["1944"]. Lloydia 7: 316.
≡Amanita watsoniana (Murrill) Murrill. 1945c ["1944"]. Lloydia 7: 327. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 276620, 345969, 291952, 284086, 291963 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes |
A. roanokensis—NCU. V. watsonianus—FLAS. | ||||||||
| revisions |
Bas. 1969. Persoonia 5: 550, figs. 361-368. Tulloss, here. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following text is derived primarily from the revision of Bas (1969), with other sources as noted. Bas (1969): Basidiomes medium to large. | ||||||||
| pileus | Bas (1969): 80 - 120 mm wide, white, with cream over disc in type, convex to planoconvex, sometimes with slight umbo, probably subviscid; context white, unchanging; margin nonstriate, appendiculate; universal veil as scattered white thin submembranous patches 4 - 12 mm wide; pileipellis peeling. | ||||||||
| lamellae | Bas (1969): adnexed to adnate, crowded to rather distant, white to pallid, narrow to moderately broad, with white subflocculose edges; lamellulae obliquely truncate to attenuate. | ||||||||
| gleba | double click in markup mode to edit. | ||||||||
| stipe | Bas (1969): 90 - 140 × 10 - 20 mm, white, subcylindric or narrowing upward, flocculose; bulb fusiform, immarginate, somewhat rooting, ca. 40 - 50 × 20 - 30 mm; context solid, white, unchanging; partial veil absent or with "vague floccose annular zone near apex"; universal veil as narrow, fragile, more or less torn limb at top of bulb against base of stipe, sometimes leaving a circlet (false partial veil) of this limb on stipe above contiguous remnants appearing to enclose bulb. | ||||||||
| odor/taste | Bas (1969): Odor "not strong of chloride of lime" (type), "faintly pleasant" (type of A. watsoniana, "like cooking meat; older like carrion or old bones" (Murrill F21875). Taste not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | Bas (1969): in two layers; suprapellis gelatinized and rather thin; filamentous hyphae 3 - 6 μm wide, yellowish in alkaline solution. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | Bas (1969): bilateral; not observable in type. | ||||||||
| subhymenium | Bas (1969): inflated ramose to subcellular. | ||||||||
| basidia | Bas (1969): 40 - 50 × 9 - 12 μm, 4-sterigmate; clamps not observed. | ||||||||
| universal veil | Bas (1969): On pileus, below surface: dingy yellowish in alkaline solution; filamentous hyphae 3 - 10 μm wide, rather abundant, interwoven; inflated cells, scattered to rather abundant, globose to ellipsoid to broadly clavate or sometimes elongate, mostly 25 - 50 μm (rarely up to 90 μm) long, terminal singly or more rarely in short chains. On pileus, exterior surface: filamentous hyphae dominant; inflated cells scarce. On stipe base: Similar to remants on pileus; but here also with inner surface layer with filamentous hyphae more abundant than in interior of limb. | ||||||||
| stipe context | Bas (1969): longitudinally acrophysalidic. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue |
Bas (1969): inflated cells mainly small, pyriform to clavate, 15 - 40 (-60) × 10 - 25 (-30) μm. [Note: Sterile.—ed.] | ||||||||
| basidiospores |
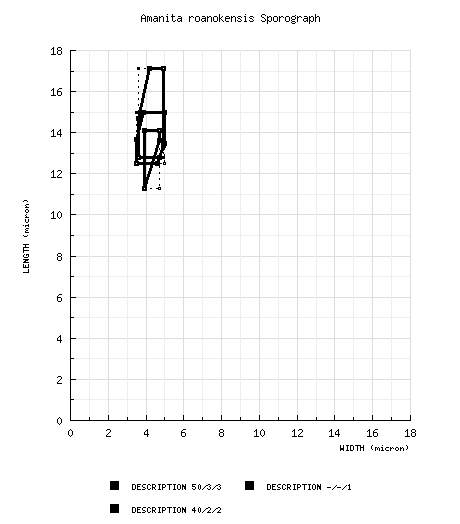
Bas (1969): [50/3/3] (11.5-) 12.5 - 15.0 × 3.5 - 5.0 μm, (Q = 2.70 - 3.90; Q = 3.20 - 3.40), yellowish, thin-walled, amyloid, cylindric to bacilliform, "sometimes slightly bent at base or slightly constricted in middle," apiculus not described; contents refractive, granular; color in deposit not recorded. "[S]ome (about 1% in types of A. roanokensis and A. watsoniana, about 50% in Murrill F 10375) with amyloid, granular warts, especially on apical half" (Bas 1969: figs. 366, 368). from type study of Amanita watsoniana by Jenkins (1979): [-/-/1] 11.3 - 14.1 (-15.6) × 3.9 - 4.7 μm, (Q = 2.90 - 4.00; Q' = 3.38), hyaline, thin-walled, amyloid, cylindric to bacilliform, often adaxially flattened; apiculus sublateral, very short, truncate; contents guttulate; color in deposit not recorded. composite of data from all material revised by RET: [40/2/2] (12.0-) 12.8 - 17.1 (-19.5) × 3.6 - 4.9 (-5.0) μm, (L = 14.4 - 14.7 μm; L' = 14.6 μm; W = 4.1 - 4.4 μm; W' = 4.2 μm; Q = (2.62-) 2.73 - 4.08 (-4.67); Q = 3.32 - 3.70; Q' = 3.51), hyaline, colorless, smooth, thin-walled, amyloid, cylindric to bacilliform, occasionally subsigmoid or subsinuate; apiculus sublateral, proportionately small; contents ??; ?? in deposit. | ||||||||
| ecology |
Bas (1969): Solitary to subgregarious. Terrestrial in dry, mixed woods. Solitary. Florida: On humus. South Carolina: In flood plain in mixed forest containing Pinus and Quercus. | ||||||||
| material examined |
Bas (1969): U.S.A.:
FLORIDA—Alachua Co. - Gainesville, 1.vii.1942 W. A. Murrill F 10357 (FLAS). Columbia Co. - Camp O'Leno, 19.x.1941 W. A. Murrill F21875 (holotype of Venenarius watsonianus, FLAS).
NORTH CAROLINA—Dare Co. - Roanoke Isl., ca. Fort Raleigh, 17.ix.1927 W. C. Coker & Braxton 8255 (holotype of A. roanokensis, NCU). from type study of Amanita watsoniana by Jenkins (1979): U.S.A.: FLORIDA— Columbia Co. - Camp O'leno, 19.x.1941 W. A. Murrill F 21875 (holotype, FLAS). U.S.A.: FLORIDA—Brevard Co. - Melbourne, 10.xi.1984 Aaron Norarevian & E. R. Yetter 4 [Tulloss 11-10-84-#4] (RET 233-6). Duval Co. - Jacksonville, 9.xi.1984 A. Norarevian & E. R. Yetter 8 [Tulloss 11-9-84-#8] (RET 234-5). Seminole Co. - Altamonte Springs, 22.vii.1958 Paul O. Schallert 7-428 (K). NORTH CAROLINA—Dare Co. - Hatteras Isl., N of Frisco, 16.vii.1981 R. E. Tulloss 7-16-81-C (RET 169-9), -D (RET 170-3), 17.vii.1981 D. C. & R. E. Tulloss 7-17-81-B (RET 169-8), -C (RET 170-6), -D (RET 169-6), -E (RET 170-2), -F (RET 170-4), -H (RET 169-7), -I (RET 170-5). SOUTH CAROLINA—Greeneville Co. - Greer, ca. border with Spartanburg Co. [34°43'30.44" N/ 82°21'32.13" W], 14.vii.2000 M. A. & R. E. Tulloss 7-14-00-A (RET 313-7). TEXAS—Hardin Co. - 6.5 km W of Silsbee, Roy Larson Sandlyand Sanctuary, 10.vi.2000 D. Pruden s.n. [Tulloss 6-10-00-J] (RET 312-3). | ||||||||
| discussion |
Bas (1969): "In the only type specimen of A. watsoniana a rather well developed volva and a basal ring [partial veil] seem to be present. A study of the tissues, however, showed that the "volva" is spurious, as it is caused by a circular splitting and partial peeling of the bulb, whereas the basal ring is formed by the [remains] of the volval limb. "The scattered, minute, adnate, amyloid warts or granules on some of the spores are thus far unique in the genus Amanita. When present, they usually occur on the apical third of the spore. Apparently they are not formed by the breaking up of an amyloid outer layer of the [spore] wall. The warts are hardly or not visible in ammonia or Congo Red." "Amanita roanokensis is characterized by very long, slender spores and a submembranous volva leaving flat, thin patches on the cap and a slight limb on top of the fusiform bulb against the base of the stem. "Probably because he applied the name A. roanokensis erroneously to forms of A. inodora, Murrill considered his A.watsoniana to be a new species. This may be inferred from his keys (1942: 1, 1948: 99) in which he placed A. watsoniana among the species with patches on the cap and A. roanokensis among those with warty or pulverulent caps. Herein the types of A.roanokensis and A. watsoniana do not differ. Thus the only differences left are the slight smell of "chloride of lime" and pure white cap of A. watsoniana and the cream cap of A. roanokensis, which in my opinion are insufficient reasons for keeping those two 'species' apart." | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.