


| name | Amanita submembranacea |
| name status | nomen acceptum |
| author | (Bon) Gröger |
| english name | "Peeling Paint Ringless Amanita" |
| images |
  1. Amanita submembranacea, Switzerland.  2. Amanita submembranacea, Norway. |
| cap |
Amanita submembranacea has a cap up to 115 mm wide; it is strongly olivaceous with a pallid margin, at least at first. Marginal striations occupy less than one fourth of the pileus radius. |
| gills |
Gills are free, not very crowded, and off-white tending to gray or brown with age; the short gills are truncate to subtruncate, plentiful, and unevenly distributed. |
| stem |
Its stem is exannulate and has a sheathing, submembranous volva at its base. This volva rapidly becomes gray after it has been split to expose the pileus, and it often has the appearance of canvas with flakes of old paint on it. |
| odor/taste | double click in markup mode to edit. |
| spores |
The spores measure (8.3-) 9.5 - 13.0 (-14.5) x (7.3-) 9.0 - 12,0 (-13.0) µm and are globose to subglobose (infrequently broadly ellipsoid) and inamyloid. Clamps are absent from bases of basidia. |
| discussion |
This species occurs in association with fir (Abies), birch (Betula), larch (Larix), or spruce (Picea), fruiting June through September. Amanita submembranacea was described from France and is now known from Norway to the Mediterranean. This species is most similar macroscopically to A. sinicoflava Tulloss, A. mortenii Knudsen & Borgen (a subarctic species the pileus of which turns medium brown or orangish medium brown after exposure to sun and drying or a few hours after collecting), A. groenlandica Bas ex Knudsen & Borgen, A. olivaceogrisea Kalaméés, and A. reidiana (which can be separated in the field by its rich dark brown pileus). At least some of the collections of "A. submembranacea" reported from England, Norway, and Scotland have proven to be A. reidiana. Also, it interesting that a somewhat similar taxon exists in South Australia (A. cheelii P. M. Kirk).—R. E. Tulloss |
| brief editors | RET |
| name | Amanita submembranacea | ||||||||||||||||||||
| author | (Bon) Gröger. 1979. Boletus 3: 27, fig. on p. 29. | ||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||
| english name | "Peeling Paint Ringless Amanita" | ||||||||||||||||||||
| synonyms |
≡Amanitopsis submembranacea Bon. 1975. Bull. Mens. Soc. Linn. Lyon 44(6): 176, fig 5. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||
| MycoBank nos. | 308592, 308606 | ||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||
| holotypes | in herb. M. Bon | ||||||||||||||||||||
| type studies | Tulloss. 1994. Mycotaxon 52: 368, figs. 39-40. | ||||||||||||||||||||
| selected illustrations |
Knudsen & Sørensen. 1980. Svampe 1: 43, fig. 2. Merlo & Traverso. 1983. Nostri Funghi Amanite: 48. Krieglsteiner. 1984. Beih. Z. Mykol. 5: fig. opposite p. 190. Outen. 1988. Mycologist 2: 34. Breitenbach and Kränzlin. 1995. Fungi Switzerland 4: 145, fig. 143. [Note there are vouchers cited.] | ||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of another researcher is based on original research by R. E. Tulloss. | ||||||||||||||||||||
| pileus | 60 - 115 mm wide, in button (with volva still covering pileus) buff over disc with slight olivaceous tint (2.5Y 8/4) and white at margin, when escaping volva darker olivaceous buff with brownish tint (2.5Y 7/4 at mid radius, between 2.5Y 6/6 and 10YR 5/4 over disc) and whitish at margin or olivaceous yellow with brownish tint (2.5Y 7/6 to 2.5Y 6/8) with whitish margin and browner disc, then bronze-brown (between 2.5Y 6/6 and 2.5Y 5/4 to 10YR 5/4, especially over disc) or more ochraceous brown (10YR 6/6) with darker bronze-brown disc (10YR 7/6) and sometimes with slightly darker zone at inner end of marginal striations and extending toward disc for about 0.10± - 0.25R and finally rather dark bronze-brown (10YR 5/8 to 10YR 7/6 and somewhat paler around umbo) or grayish brown with hardly any olive-buff tint remaining (10YR 5/3) and darker over disc with fulvous tint (10YR 4/4 to 7.5YR 4/4) or umber with very slight fulvous tint (7.5YR 5/2-4) or rather olivaceous brown with coppery tint or slightly bronzy [Séguy 337 (redder than 5YR 6/6) mixed with Séguy 301 (deep olive with brownish tint)] and with paler margin slightly more yellowish [Séguy 338 (pale brown)], sometimes in old pilei paler [slightly brownish buff to silvery brownish buff (2.5Y 7/6)], hemispherical to evenly convex with slight umbo especially visible after full expansion to conico-convex, then plano-convex, finally planar with umbo or plano-concave with umbo, sometimes depressed around umbo, smooth, viscid to subviscid to sticky, lustrous, not innately fibrillose; context unrecorded; margin long striate (0.25 - 0.5±R), even, slightly inflexed or decurved at first, finally flaring upward, sometimes splitting; universal veil absent or in irregular, subpulverulent, grayish patches, mostly concentrated over disc. | ||||||||||||||||||||
| lamellae | free, often with decurrent lines at apex, subdistant, not very crowded to moderately crowded to crowded, white or white with creamy reflection in mass, becoming sordid whitish and then slightly brownish or browning with age, drying pale brownish yellow, 10 - 13 mm broad, eventually subventricose, with entire subflocculose edge rather straight at first, with flocculence white to distinctly grayish brown; lamellulae truncate to subtruncate to concavely truncate, unevenly distributed, plentiful to moderately scarce to scarce. | ||||||||||||||||||||
| stipe | 85 - 180 × 6 - 17 ( 24) mm, subcylindric to narrowing upward, slightly bulbous at base, decorated with minute thin white flocculence at first, with flocculence sometimes clearly terminating at line below top of (hidden by) volval limb, with ground soon tinted pale pinkish ochraceous to pale salmon to salmon ochraceous (10YR 7/6 with touch of 7.5YR 7/6) under paler floccosity, sometimes with flocculose layer giving rise to flocculose squamules at its surface and tending to become pale pinkish ochraceous cream to pale or very pale salmon, with floccosity developing into appressed pale grayish buff fibrils, finally with rather dark gray-brown to fulvous-ochraceous brown appressed fibrils in zigzag pattern on paler grayish brown to beige (near apex) ground or (as in protologue) smooth and white at apex, somewhat pulverulent below this, and progressively decorated toward base with grayish yellow rings or zigzag (snakeskin) pattern; context white to whitish to slightly creamy to grayish (in very dark specimens), near surface sometimes concolorous with surface, unchanging, stuffed at first, then partially or entirely hollow, with central cylinder of diameter often greater than 50% of stipe diameter and extending down into slightly swollen base; exannulate; universal veil submembranous to felted-membranous, sometimes flaring widely above point of attachment to stipe, tending to crumble easily or become irregular torn, with outer surface rather felted flocculose, sometimes with surface breaking up into thin plaques suggestion paint peeling from an old canvas (visible is some exsiccata), at times suggesting false annulus near stipe base by being in one or several separated zones of patches, at times incomplete, derived from often limbate volva with 2 - 5 lobes, rarely complete and saccate (then rather ample), 35± - 55± mm from stipe base to highest point on limb, rather thick, at first pallid evently brownish gray at upper edge and on exterior surface grading through shades of gray (e.g., grayish buff to pale gray to slightly pinkish gray to slightly sordid buff) to whitish at base, interior surface usually buff gray (10YR 7/2-3 to 10YR 6/3-4), relatively appressed to stipe base for bottom 10 - 30 mm, at times slightly strangulate above this point, with interior longitudinally pleated or grooved in luxuriant specimens, often at first with rather pronounced flocculose ridge on stipe 4 - 10 mm above attachment of volva. | ||||||||||||||||||||
| odor/taste | Odor mild or lacking. Taste mild and somewhat nutlike to indistinct, with delicate flavor when cooked with butter (Bas 7021). | ||||||||||||||||||||
| macrochemical tests |
Phenol - sordid reddish brown to pinkish wine colored, not bright cassis, but between reactions in A. fulva and in A. crocea. Phenolaniline - on context of stipe base, within 1 min. vinaceous pink, becoming deep vinaceous red after 5± min., then slowly becoming purple-brown. Test vouchers: holotype and Bas 7021. | ||||||||||||||||||||
| pileipellis | 50 - 95 µm thick, gelatinizing just at surface or to depth of up to 30 µm, colorless in upper half, orange-brown in lower half, with pigmentation extending slightly into immediately adjacent context; filamentous, undifferentiated hyphae 1.5 - 6.2 ( 9.4) µm wide, densely packed, interwoven, subradially arranged; vascular hyphae 3.5 - 8.4 µm wide, uncommon to locally moderately common. | ||||||||||||||||||||
| pileus context | filamentous, undifferentiated hyphae 2.5 - 15.0 µm wide, branching, plentiful, with those of smaller diameter mostly in loosely interweaving fascicles, with those of largest diameter not in fascicles, with some branched elements of largest diameters; acrophysalides dominating, terminal, thin-walled, subfusiform to clavate to broadly clavate to elongate to ovoid to ellipsoid, up to 175 × 60 µm; vascular hyphae 4.2 - 16.0 µm wide, branching, infrequent, locally in tangles | ||||||||||||||||||||
| lamella trama | bilateral, largely collapsed in isotype; with angle of divergence negligible?? to 90°??, mostly at extremes of this range??; consisting of central stratum (wcs = 35 - 50 µm in isotype) flanked on each side by a region up to 20 µm wide containing occasional terminal inflated cells (sometimes several together) in a single (rarely double, more frequently sparsely populated to locally absent) layer; filamentous, undifferentiated hyphae 1.8 - 9.5 µm wide, branching, closely packed, largely co-parallel, branched cells and inflated tip segments up to 29 µm wide; inflated cells (largely collapsed in isotype) intercalary, ellipsoid to broadly ellipsoid to subglobose to irregular, up to 118 × 41 µm, with majority less than 70 µm long, thin-walled [, not or minimally diverging??]; vascular hyphae 3.0 - 7.5 µm wide, scarce. | ||||||||||||||||||||
| subhymenium | largely collapsed in isotype; comprising branching hyphae having very short segments and small inflated cells and subcoralloid elements, with basidia arising from short uninflated hyphal segments or small inflated cells. | ||||||||||||||||||||
| basidia | 32 - 82 × 8.8 - 19.5 µm, thin-walled, predominantly 4-sterigmate, occa¬sionally 2-sterigmate, with sterigmata up to 6.0 × 3.2 µm; clamps rare or absent. | ||||||||||||||||||||
| universal veil | On pileus: absent. At stipe base, exterior surface: as occasionally disrupted layer of partially gelatinized to gelatinized fascicles of hyphae one or two fascicles thick, commonly closely packed, but also somewhat loosely interwoven in some regions, dominantly sublongitudinally oriented, with somewhat gelatinized tissue of interior exposed in some areas; filamentous, undifferentiated hyphae 1.0 - 8.5 µm wide, branching, almost entirely in aforementioned fascicles; vascular hyphae 2.5 - 4.8 µm wide, branching, not included in fascicles. At stipe base, interior: all elements with slightly brownish tint in region near exterior surface; filamentous, undifferentiated hyphae 3.0 - 5.8 µm wide, plentiful, branching, loosely interwoven, in fascicles or not; inflated cells dominating, often very densely packed, terminal singly or in short chains, with walls thin or infrequently up to 0.5 µm thick, broadly fusiform to elongate to narrowly clavate to broadly clavate to subpyriform to subglobose, infrequently branched, up to 116 × 67 µm, with tendency to take on more elongate forms near surfaces; vascular hyphae 1.8 - 5.6 µm wide, scarce; clamps not observed. At stipe base, inner surface: like interior, but with proportionately more hyphae and with inflated cells on average smaller and more elongate, somewhat gelatinized. | ||||||||||||||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 0.7 - 9.8 µm wide, dominating; acrophysalides common, thin-walled, up to 307 × 63 µm; vascular hyphae 3.5 - 18.2 µm wide, common to plentiful, branching, occasionally in loose tangles; clamps not observed. | ||||||||||||||||||||
| basidiospores |
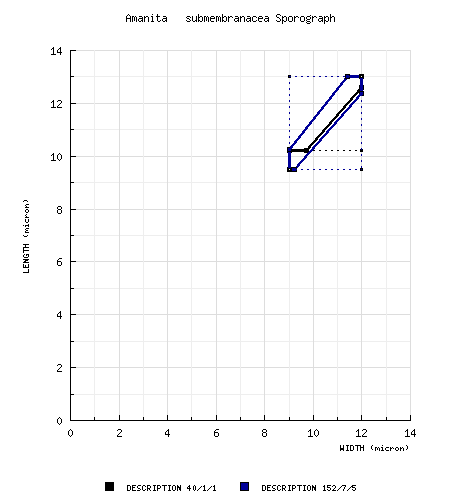
Tulloss (1994): [40/1/1] (9.2-) 10.2 - 13.0 (-13.8) × (8.2-) 9.0 - 12.0 (-12.5) μm, (L = 11.5 μm; W = 10.6 μm; Q = (1.04-) 1.05 - 1.14 (-1.22); Q = 1.09). composite data from all material revised by RET: [152/7/5] (8.3‑) 9.5 - 13.0 (‑14.5) × (7.3‑) 9.0 - 12.0 (‑13.0) µm, (L = (9.8‑) 10.3 - 12.3 µm; L’ = 11.2 µm; W = (9.2‑) 9.6 - 11.3 µm; W’ = 10.3 µm; Q = (1.0-) 1.03 - 1.14 (‑1.22); Q = (1.06‑) 1.07 - 1.10; Q’ = 1.08), inamyloid, thin-walled, smooth, hyaline, colorless, globose to subglobose to broadly ellipsoid, usually at least slightly adaxially flattened; apiculus rather large, abrupt, subcylindric to cylindric; contents mono- or multiguttulate; off-white in deposit. | ||||||||||||||||||||
| ecology | Solitary to subgregarious, at times abundant. France: At 1100 m elev. In Abies forest with admixture of Betula in low vegetation and mosses under Vaccinium or with Picea in acid soil. Germany: At 625 m elev. Over acid multicolor sandstone, in grassy clearing of of submontane forest under Picea abies and Abies albs. Italy: At 1400± m elev. Under Picea abies and Larix decidua or in Picea-Larix forest with much Vaccinum myrlitus on acid soil. Switzerland: In Pinus cembra forest in deep humus over acid rock with scattered Picea and Larix and much V. myrlitus and Lusula and Arnica montana. | ||||||||||||||||||||
| material examined | FRANCE: ARDENNES—Croix (front Belgique) , 30.ix.1978 M. Bon & S.M.N.F. [Bon 780930] (in herb. M. Bon=>LIP). HAUTE-LOIRE—Fontanet, ca. Le Puy, 17.ix.1974 M. Bon 740917 (holotype, in herb. M. Bon=>LIP, n.v.; isotype, L, fragments). GERMANY: BADEN WÜRTTEMBERG—Bad Liebenzell-Maisenbach, "Rohrbrunnen" [625 m], 9.ix.1998 Andreas Gminder 98/338 (in herb. A. Gminder, RET 306-4, nrITS-LSU seq'd.). Homburg, westlich Niederwasser, Nähe Gasthof, "Schöne Aussicht," 3.x.1997 W. Pätzold 97/426 (in herb. Gminder; RET 305-6, nrITS & nrLSU seq'd.). ITALY: TRENTINO—Frassilongo, Prati Imperiali, 29.vi.1997 Marco Floriani s.n. (RET 270-6). Valsugana, Cinque Valli between Roncegno and Vetriolo Terme, 3.ix.1987 C. Bas 9144 (L; RET 270-6). POLAND: WOJEWÓDZTWO MALOPOLSKIE—Tatra County - Hohe Tatra, Zakopane, trail to Trzedniowanski Wierch [1450 m], 28.ix.1992 Andreas Gminder 95/446 (in herb. Gminder; RET 306-6, nrITS-LSU seq'd.). SPAIN: UNKN. PROV., unkn. loc., 12.x.2019 Paco Villalongo PV19101202 (RET 884-1, nrITS seq'd.). SWITZERLAND: GRAUBÜNDEN—3 km E of St. Moritz, God da Staz hill, 26.viii.1976 C. Bas 7021 (L; RET 002-1). | ||||||||||||||||||||
| discussion |
The macroscopic description given here benefits greatly from extensive notes of Dr. C. Bas on the Valsugana and St. Moritz collections. This anatomical description differs from that of Tulloss (1988) in several points. My earlier description relied heavily upon material other than the holotype, which was not examined in detail at the time and of which, apparently, very little remains. After the study of the holotype reported above, I regard the earlier description as in error in reporting the small, subglobose to broadly ellipsoid, terminal, inflated cells in the subhymenial region as part of the subhymenium rather than part of the subhymenial base. In the previous description the pileipellis is described as much thicker than was found in the holotype and larger, clavate, inflated cells are described in the lamella trama. In addition to these differences, new data has been added. The reader may wish to compare the present species with the following: A. groenlandica, A. mortenii, and A. reidiana. In the U.K., material previously determined as A. submembranacea has been redetermined as A. reidiana upon revision by RET. | ||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||
| editors | RET | ||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita submembranacea |
| bottom links | [ Keys & Checklist/Picturebooks ] |
| name | Amanita submembranacea |
| bottom links | [ Keys & Checklist/Picturebooks ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.