


| name | Amanita romagnesiana |
| name status | nomen acceptum |
| author | Tulloss |
| english name | "Romagnesi's Ringless Amanita" |
| synonyms |
≡Amanita crocea var. aurantiofulva (E.-J. Gilbert) Romagn. |
| cap |
The cap of Amanita romagnesiana is up to 120 mm or more wide, rounded-conic to campanulate at first, becoming obtuse, lacking an umbo, with a short-striate margin in the dried specimen. The original description states that the cap is bright orange-tawny, occasionally with some regions a little more brownish orange. In some apparently contaxic collections, the pileus is described as somewhat paler than this. The volva is absent. |
| gills |
The gills are crowded, pale cream, and rather narrow in dried specimens. The short gills are truncate (?), very infrequent, and unevenly distributed. |
| stem |
The stem is up to 300 × 15 - 20 mm, pale brownish orange, narrowing upward slightly, usually decorated with pale orange floccules, and exannulate. The flesh is white or very pale brownish orange near the surface. The saccate volva is white in the exterior, pale orangish brown in the interior, elongate, often strangulate, and membranous. |
| spores |
The spores measure (8.0-) 9.4 - 12.8 (-21.0) × (7.2-) 7.9 - 11.4 (-17.0) µm and are subglobose to broadly ellipsoid (rarely ellipsoid) and inamyloid. Clamps are not found at bases of basidia. |
| discussion |
Amanita romagnesiana was originally described from France (Dep. Manche) from deciduous forest or mixed forest containing Norway Spruce. Herbarium collections have been located that orginated in Belgium and Germany. This is species is related to species in the group around A. crocea (Quél. in Bourd.) Singer ex Singer and has some similarities to the group including A. fulva (Schaeff.) Fr.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita romagnesiana | ||||||||
| author | Tulloss. 2000. Mycotaxon 75: 329. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Romagnesi's Ringless Amanita" | ||||||||
| synonyms |
≡Amanita crocea var. aurantiofulva (E.-J. Gilbert) Romagn. 1992. Bull. Trimestriel Soc. Mycol. France 108: 80, 86.
≡Amanita vaginata [var. fulva] f. aurantiofulva E.-J. Gilbert. 1918. Gen. Amanita Pers.: 145.
≡Amanita vaginata [subsp. fulva] f. aurantiofulva (E.-J. Gilbert) Konrad & Maubl. 1924. Icones Selectae Fungorum 6: 33.
[Note: E.-J. Gilbert (1941. Iconogr. Mycol. (Milan) 27, suppl. (2): 212) treated the present taxon as a synonym of Amanita crocea.] The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| etymology |
Romagnesi + -ana, suffix indicating possession; hence, "of Romagnesi" Honoring Henri Romagnesi. | ||||||||
| MycoBank nos. | 467531 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| neotypes | in herb. H. Romagnesi (PC) | ||||||||
| neotypifications | Romagnesi. 1992. Bull. Trimestriel Soc. Mycol. France 108: 86. | ||||||||
| type studies | Tulloss, here. | ||||||||
| selected illustrations | Romagnesi. 1961. Nouv. Atlas Champ. 3: pl. 178. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is based entirely on the protolog of the present taxon and the type study of Tulloss (1994). | ||||||||
| pileus | up to 120 or more mm wide, bright orange-tawny, occasionally with some regions a little more brownish orange, with color over the disc barely more saturated than elsewhere, rounded-conic to campanulate at first, becoming obtuse, lacking an umbo; context not described; margin short striate in exsiccatum [length when fresh not estimatable from illustration]; universal veil absent; pileipellis easily peeling at mid-radius in exsiccatum. | ||||||||
| lamellae | crowded, pale cream, rather narrow in exsiccatum; lamellulae [based on exsiccatum] truncate(?), very infrequent, unevenly distributed, with all observed having length roughly 0.5R. | ||||||||
| stipe | up to 300 × 15 - 20 mm, pale brownish orange (pale rufous), narrowing upward slightly, usually decorated with pale orange floccules, less often intensely pruinose; context white or very pale brownish orange near the surface; exannulate; universal veil as saccate volva, with white exterior and pale orangish brown interior, elongate, with highest point on limb up to 40 - 50 mm from stipe base, weakly adnate for about half of height, then expanding in a broad, funnel-shaped form, membranous, but sometimes weakly so (on exsiccatum, two large patches distributed up stipe for roughly 3 cm above remainder of volval limb), sometimes with a strangulate region below limbus internus or having entire limb above limbus internus separated entirely from remainder of sac and elevated on stipe (Romagnesi, 1961, 3: pl. 178), with limbus internus brief and relatively thick and placed at point of attachment to stipe. | ||||||||
| odor/taste | Odor and taste not described. | ||||||||
| macrochemical tests |
none described. | ||||||||
| pileipellis | 210 - 220 µm thick, orangish yellow, fully gelatinized only at surface; filamentous, undifferentiated hyphae 2.2 - 18.0 µm wide, branching, densely packed, dominantly subradially oriented, but often criss-crossing, partially gelatinized or ungelatinized below surface, occasionally fragmented, often with yellowish (occasionally subrefractive) walls; vascular hyphae not observed. | ||||||||
| pileus context | filamentous, undifferentiated hyphae 2.5 - 10.0 µm wide, branching, plentiful, singly and in fascicles, interwoven in loose lattice; acrophysalides broadly clavate to clavate to fusiform, up to 134 × 35 µm, thin-walled; vascular hyphae not observed. | ||||||||
| lamella trama | bilateral, with shallow angle of divergence; wcs = (25-) 30 - 45 µm; subhymenial base comprising branching hyphae and inflated cells, with intercalary inflated cells clavate to ovoid to ellipsoid (up to 45 × 19.0 µm) and often with slightly thickened walls, with some inflated cells extending into subhymenium (and these with major diameter nearly perpendicular to central stratum; filamentous, undifferentiated hyphae 2.8 - 8.0 µm wide, branching, with some intercalary segments in central stratum partially inflated (up to 16.0 µm wide); terminal, divergent inflated cells not observed; vascular hyphae not observed. | ||||||||
| subhymenium | wst-near = (25-) 30 - 60 µm; wst-far = 55 - 80 µm; comprising inflated cells (broadly clavate to ovoid to ellipsoid to subglobose to irregular, up to 22 × 15.5 µm) and partially inflated and uninflated short hyphal segments, with largest inflated cells tending to be toward subhymenial base, with basidia arising from cells of all types, with 20 - 35 µm between subhymenial base and nearest base of basidium/-ole, with 45 - 55 µm between subhymenial base and farthest base of basidium/-ole. | ||||||||
| basidia | 51 - 70 × 12.5 - 15.0 µm, both 4- and 2-sterigmate; clamps not observed. | ||||||||
| universal veil | On pileus: absent. On stipe base, exterior surface: loosely interwoven layer of broad fascicles of filamentous, undifferentiated hyphae (1.8 - 8.8 µm wide, branching, often partially gelatinized) with large gaps through which interior tissue visible; vascular hyphae 4.0 - 8.0 µm wide, as partially gelatinized scarce fragments. On stipe base, interior: filamentous, undifferentiated hyphae 1.2 - 10.5 µm wide, plentiful to dominating, branching, singly and in broad fascicles loosely interwoven in open lattice, occasionally partially gelatinized; inflated cells plentiful to locally dominating, thin-walled, globose to subglobose to subpyriform to broadly ellipsoid (up to 74 × 69 µm), ellipsoid to ovoid to broadly clavate (up to 80 × 52 µm), clavate to narrowly clavate to fusiform (95 × 39 µm); vascular hyphae not observed. On stipe base, inner surface: very thin layer of densely packed, dominantly sublongitudinally oriented, filamentous, undifferentiated hyphae (1.4 - 8.4 µm wide, branching, collapsed, partially to totally gelatinized) through which interior tissue visible; vascular hyphae 4.6 - 13.3 µm wide, branching, as very few scattered fragments, partially gelatinized or not. | ||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 5.1 - 15.4 µm wide, branching, plentiful to locally dominant near surface; acrophysalides thin-walled, plentiful to locally dominant in interior, up to 375 × 40 µm; vascular hyphae 3.2 - 15.4 µm wide, rather common, sinuous, loosely coiling, locally knotted or tangled, locally having abrupt constrictions. | ||||||||
| lamella edge tissue | widely scattered remains of layer up to 40 µm thick; cells scant (specimen just maturing when dried), usually in one layer, sometimes two layers, up to 40 × 25 µm, partially gelatinized, with one or two strands of filamentous, undifferentiated hyphae 3.5 - 5.6 µm wide and radially oriented. | ||||||||
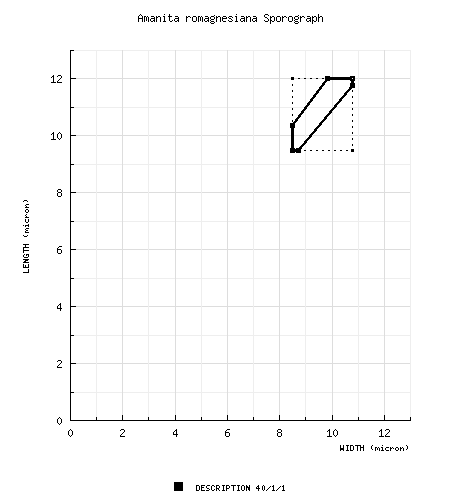
| basidiospores | [40/1/1] (8.0-) 9.5 - 12.0 (-16.0) × (7.5-) 8.5 - 10.8 ( 14.2) µm, (L = 10.8 µm; W = 9.5 µm; Q = (1.07-) 1.09 - 1.22 (-1.31); Q = 1.14), hyaline, colorless, smooth, thin-walled, inamyloid, subglobose to broadly ellipsoid, rarely ellipsoid, adaxially flattened, infrequently expanded at one end; apiculus sublateral, cylindric, occasionally quite large; contents monoguttulate; white in deposit. | ||||||||
| ecology | In deciduous woods or in mixed woods including Picea abies L. | ||||||||
| material examined |
FRANCE: MANCHE—Cotentin peninsula, Forêt de Saint-Sauveur-le-Vicomte, 12.xi.1976 H. Romagnesi 76.117 | ||||||||
| discussion |
The single specimen of the neotype collection has about an even number of 4-sterigmate and 2-sterigmate basidia in the mid-region of the lamellae. Spores are plentiful in all mounts, but there are regions of the hymenium with few mature basidia. The apparent youth of the material when dried suggests that more mature material will have basidia dominantly 4-sterigmate. In Malençon's painting (Romagnesi, 1961: pl. 178), the marginal striations of the pileus are shown to be extremely short, which character is confirmed in the exsiccatum examined. Also, the pileus has a rather ruddy tone; and the volval sac is elevated with a distinctly strangulate zone remarked upon in the accompanying text. Romagnesi describes the spores as having dimensions from which one would estimate a Q’ of 1.20 - 1.25; however, this is not the case in the spores measured from the holotype. Romagnesi (1992) explicitly rejects the text accompanying Malençon's painting (Romagnesi, 1961) saying that this text is a compilation of descriptions of A. crocea and the present taxon. He further states that the painting represents material collected in the Tyrol by Malençon, possibly in the company of M. Moser, and that such material might have been deposited in IB or MPU. In A. crocea, there are hyphal segments running sybparallel to the central stratum throughout the subhymenial tree. The subhymenial base is largely composed of narrow, intercalary, subfusiform cells and uninflated hyphal segments. There is an occasional broader inflated cell. The cells of the subhymenium are smaller and usually less fully inflated. The interior layer of the universal veil limb comprises filamentous, undifferentiated hyphae 2.1 - 11.6 µm wide, with occasional vascular hyphae and fewer inflated cells than are found in the same tissue of A. crocea var. aurantiofulva. In my opinion, the two “varieties” differ too much to be assigned to a common species. The pileipellis of A. crocea is 100 - 160 ( 240) µm thick. Along the lamella edge, there is an extensively preserved layer of inflated cells (up to 58 × 38 µm) up to 163 µm thick (5 to 7± cells thick) when ungelatinized and up to 80 µm thick when partially gelatinized. This layer may contribute to the decoration of the stipe of A. crocea. I was able to examine Tulloss 9-4-88-I (from Scotland, U.K.) in fresh condition. The limbs of the volva were leathery and rather thick; the pileus color was a rather pale yellowish orange (more orange than 4A4, more yellow than 10YR 8/6); and the stipe was pale orangish and shaggy as is typical of A. crocea with densely set, upward pointing, pallid, fibrillose squamules that became dark orange on the tips or (when handled) more extensively. A. crocea has vascular hyphae in its pileipellis and less frequent inflated cells in the interior of its universal veil. Based on the limited number of specimens examined, Amanita subnudipes differs from the present taxon in the following characters: Unfortunately, the holotype of A. subnudipes is becoming damaged by the attack of a mold; and the lamella trama could not be rehydrated at all. Therefore, the judgments behind the above list are in part based on non-type material. The two items preceded by question marks are those that seem to me to be mostly likely to be influenced by inadequate sample size. Nevertheless, it is clear from the above that the two entities are not conspecific. Despite an apparent relationship to A. fulva (Tulloss, unpub. res.), the color of the pileus; the short marginal striations of the pileus; spores with Q > 1.10; a considerably narrower central stratum (compared to wcs = 85 - 90 µm for A. fulva); and a subhymenial tree apparently lacking in filamentous, undifferentiated hyphae running roughly parallel to the central stratum separate A. crocea var. aurantiofulva from A. fulva at the rank of species. Amanitopsis vaginata var. cinnabarina Killerman [1931. Denkschr. Bayer. Bot. Ges. Regensburg 18 (neue Folge 12): 11] was suggested as a synonym of A. crocea by Fraiture (1993). Considering the very sparse protolog of the Killerman taxon (especially considering the pileus color reflected in the varietal epithet), it is more likely that Amanitopsis vaginata var. cinnabarina would be a synonym of Amanita crocea var. aurantiofulva than of the type variety or A. subnudipes. I would be very interested in hearing from any reader who knows the whereabouts of a Killerman collection of his var. cinnabarina. The packet loaned to me and marked “neotype” by prof. Romagnesi is numbered “76-197” in script and is glued to a sheet on which “197” is typed in the upper left hand corner. In addition, a cardboard tab is attached to the sheet on which the number “76-197” was written and is corrected in a darker ink to read “76-117.” Due to fading of the ink, the handwritten number on the packet might be mistaken for “76-117.” The latter was the collection number cited when the neotype was designated (Romagnesi, 1992). | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita romagnesiana |
| bottom links | [ Keys & Checklists ] |
| name | Amanita romagnesiana |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.