


| name | Amanita subalbida |
| name status | nomen acceptum |
| author | Cleland |
| intro | The following is based on the description by Bas (1969). |
| cap | The cap of Amanita subalbida is about 30 - 45 mm wide, somewhat irregularly convex to plane with a slightly depressed center, dry, pallid brownish to nearly white, with a nonsulcate, slightly appendiculate margin. The cap is covered with thin, pulverulent-subfelted, concolorous, crust-like remnants of volva especially over the center. |
| gills | The gills are crowded, slightly adnexed, moderately broad, and white. The short gills are truncate to rounded-truncate. |
| stem | The stem is about 30 - 50 × 8 - 10 mm, equal, solid, white, and without any remnants of volva. The stipe's basal bulb is subglobose to ellipsoid and 8 - 15 mm wide. |
| odor/taste | The odor and taste of this mushroom are unknown. |
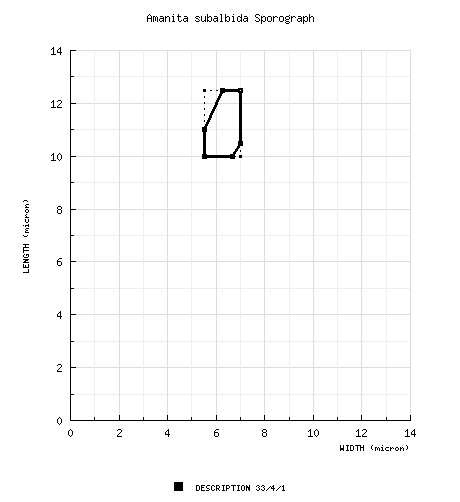
| spores | The spores measure 10 - 12.5 (-13.5) × 5.5 - 7 (-8) µm and are amyloid and ellipsoid to elongate. Clamps are present, but rare, at bases of basidia. |
| discussion |
Amanita subalbida was originally described from South Australia, and Bas knew it only from the type. Bas placed this species in his stirps Grossa (see A. grossa (Berk.) Sacc.).—R. E. Tulloss |
| brief editors | RET |
| name | Amanita subalbida | ||||||||
| author | Cleland. 1931. Trans. Roy. Soc. S. Austral. 55: 152. | ||||||||
| name status | nomen acceptum | ||||||||
| synonyms |
≡Aspidella subalbida (Cleland) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 79, tab. 52 (fig. 3). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 279089, 284334 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | AD | ||||||||
| revisions |
Bas. 1969. Persoonia 5: 510, figs. 291-293. Reid. 1980. Austral. J. Bot., Suppl. Ser. 8: 56, 40(a-c), 93. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The text below is derived from the revision of Bas (1969). from Bas (1969): Basidiome small. | ||||||||
| pileus | from Bas (1969): 30 -45 mm wide, pallid brownish to nearly white, somewhat irregularly convex to planar, with slightly depressed center, dry ("judging by the microscopical structure of the pileipellis, probably viscid when moist"); context not described; margin nonsulcate, slightly appendiculate; universal veil in crust-like fragments most often over disc, subfelted, concolorous(?), thin. | ||||||||
| lamellae | from Bas (1969): slightly adnexed, crowded, slightly adnexed, white, ventricose, moderately broad, with flocculose edge when young; lamellulae truncate to rounded truncate. | ||||||||
| stipe | from Bas (1969): ca. 30 - 50 × 8 - 10 mm, cylindric, subpulverulent-subfibrillose; bulb rather abrupt, globose to ellipsoid, ca. 8 - 15 mm wide; context solid; partial veil apical, thin, pendent, subfelted-membranous, vaguely striate [above], sometimes fugacious; universal veil absent or as slight felted rim at top of bulb. | ||||||||
| odor/taste | not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | from Bas (1969): rather thick, gelatinized near surface, pale brownish in alkaline solution; filamentous hyphae 2.5 - 5 μm wide, interwoven; vascular not described. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | from Bas (1969): bilateral; clamps rare. | ||||||||
| subhymenium | from Bas (1969): ramose to subcellular. | ||||||||
| basidia | from Bas (1969): 40 - 50 × 10 - 12 μm, 4-sterigmate; clamps rare. | ||||||||
| universal veil | from Bas (1969): On pileus: pale brownish in alkaline solution, with elements irregularly disposed; filamentous hyphae 2.5 - 8 μm wide, loosely interwoven; inflated cells scattered, globose or pyriform or clavate or elongate, up to 100 × 70 μm], terminal singly or in short chains; vascular hyphae not described. On stipe: with outer part of rim comprising tissue like that on pileus, but with smaller inflated elements. | ||||||||
| stipe context | from Bas (1969): longitudinally acrophysalidic; filamentous hyphae scattered; acrophysalides "usually not longer than 200 μm"; vascular hyphae not described. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | from Bas (1969): filamentous hyphae 1.5 - 4 μm wide, loosely interwoven; inflated cells pyriform to clavate, 10 - 35 × 8 - 20 μm. | ||||||||
| basidiospores |
from type study of Bas (1969): [33/4/1] 10.0 - 12.5 (-13.5) × 5.5 - 7.0 (-8.0) μm, (Q = 1.5 - 2.0; Q = 1.65 - 1.80), colorless, hyaline, thin-walled, amyloid, ellipsoid to elongate, sometimes slightly attenuate toward the base or substrangulate; apiculus sublateral (per figure); contents "often" refractive; color in deposit not recorded. from type study of Grgurinovic (1997): [50/-/1] 9.6 - 14.3 (-16.4) × 5.9 - 8.0 μm, (L' = 11.6 μm; W' = 6.6 μm; Q' = 1.70), amyloid, elongate. [Note: We have not estimated a range for Q based on the (Grgurinovic 1997) spore data for this species in order to prevent automatic generation of a sporograph that we believe would be misleading. We evaluated the Amanita spore length and width ranges from (Grgurinovic 1997) in comparison to comparable data published by other authors and often based on revision of the same specimens. This experiment involved a total of 19 descriptions of a total of 13 species from the work of 3 authors. In a range of the form "x - y" of spore length (width) from (Grgurinovic 1997) compared to a range of the form (a-) b - c (-d) of spore length (width) in the other works, the value of "y" was compared to the value of "c" as a ratio. In the case of spore length ranges, on average (per author), the ratio y/c ranged from 1.06 - 1.10 (possibly due to the non-segregation of a "d" value in the ranges of concern). In the case of spore width ranges, on average (per author), the ratio ranged from 1.14 - 1.23 (indicating the probability of compounding causes at play—possibly, the absence of the "d" value in the ranges of concern and failure to restrict spore measurement to spores strictly presenting in lateral view). When sporographs were attempted from the Grgurinovic data, the results were not useful.—ed.] | ||||||||
| ecology |
from protolog: "Half-buried in sandy soil." from type study of Bas (1969): In small group (per number of basidiomes in type collection—4). | ||||||||
| material examined | from type studies of Bas (1969), Reid (1980), and Grgurinovic (1997): AUSTRALIA: SOUTH AUSTRALIA—Sturt Co. - Kinchina, 8.vi.1925 J. B. Cleland s.n. (holotype, ADW 9267 => AD 3133). [Note: accession numbers were obtained from (Grgurinovic 1997).] | ||||||||
| discussion |
from Bas (1969): "The colour of the dried materiall is remarkably dark; the caps and gills are dark chocolate brown, but the remnants of the volva on the cap are pale ochraceous buff, and the stems and bulbs pale ochraceous brown with a slight purplish tinge. "Clamps are distinctly present but scarce. It is my impression that they are to be found especially at the base of the first formed basidia. [It was later Bas' opinion that the most likely place to find clamps in a given species was in the part of the gill that produces mature basidia last—that portion farthest from the stipe and nearest the gills bottom edge.—ed.] "One of the four type specimens has a slight volval rim at the top of the bulb. This, together withthe presence of clamps, could be an indication that A. subalbida should be placed in stirps Limbatula of subsection Limbatulae, but since in the protologue the volval remains on the cap are described as mealy and the rim at the base of the stem as friable, I have placed it in stirps Grossa. "Amanita straminea, a small species in stirps Straminea, bears some resemblance to A. subalbida. It differs, however, in that it lacks clamps, has straw yellow gills and lacks volval remnants at the base of the stem." The reader may wish to compare this species with A. angustispora, which seems very similar in many respects. Reid (1980: 57) describes briefly two collections that he suggests are assignable to the present species. He emphasizes spore size in both cases; however, the sporographs representing his spore data (with estimated values of Q) are essentially or completely disjunct from the sporograph (above) based on Bas' data. Both taxa are said to lack a skirt-like partial veil; and in one case, it is explicitly stated that there are no remnants of universal veil on the stipe's bulb. Under these circumstances, the editor has decided not to include the two collections involved here without further information confirming the placement. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.