


| name | Amanita oblongispora |
| name status | nomen acceptum |
| author | Contu ex Tulloss & Contu |
| english name | "Contu's Ringless Amanita" |
| images |
 1. Amanita oblongispora, Sardinia, Italy. |
| cap |
The cap of A. oblongispora is 65 - 80 mm wide, campanulate at first, later planoconvex to planar, often with a prominent and broad umbo, glabrous, moist and lubricious at first to slightly viscid, then dry, with a striate-sulcate margin (up to 7 mm or 10 - 20% of the radius). The cap is light pearl gray, with disc brown-ocher, darkening with age. The volva is absent or as a large patch or patches, submembranous to membranous, rather thick, white to off-white. |
| gills |
The gills are free, subdistant, white, orangish tan to grayish brown to sordid tan in exsiccata, rather broad and thick. Short gills are present. |
| stem |
The stem is 80 - 150 × 6 -15 mm, cylindric to narrowing upward, subsmooth, white to whitish above, exannulate, sometimes with rather pale brown-bister fibrils in bands below. The flesh is white to slight bister towards base, stuffed to hollow, quite firm, but fragile and quick to decay. The small, cup-like volva or fragile, saccate volva is white to ochraceous, friable to submembranous, extending up to 30 mm up stem, not easily removed from substrate. |
| spores |
The spores measure (8.2-) 10.5 - 13.8 (-16.2) × (7.2-) 7.8 - 10.8 (-13.5) µm and are inamyloid and broadly ellipsoid to ellipsoid (occasionally elongate, infrequently cylindric). Clamps are rather common at bases of the basidia. |
| discussion |
This species was originally described from Italy (Sardinia) and is still known only from this region. This species seems to be rather isolated if my observation of clamped basidia is correct. Taxa with broadly ellipsoid to ellipsoid spores and weak or relatively weak volvas include taxa similar to A. malleata (Piane ex Bon) Contu on the one hand and taxa similar to A. mairei Foley on the other. At the moment, I believe the relationship of A. oblongispora is closer to the first of these groups; however, this observation still fails to place A. oblongispora definitively. Obviously, work remains to be done in these groups.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita oblongispora | ||||||||
| author | ("oblongospora") Contu ex Tulloss & Contu in Tulloss. 1994. Mycotaxon 52: 358, figs. 33-34. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Contu's Ringless Amanita" | ||||||||
| synonyms |
≡Amanita oblongospora Contu nom. inval. 1988. Bol. Soc. Micol. Madrid 12: 90. [All requirements for valid publication were met except that a type was not specified, although the herbarium in which it is deposited was mentioned. ICBN §37.1, §37.3.] The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| etymology | oblongus "oblong" + spora "spore"; hence, "oblong spore" | ||||||||
| MycoBank nos. | 362537 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | CAG | ||||||||
| selected illustrations | Consiglio. 2000. Boll. Gruppo Micol. G. Bresadola 43(2): 225. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present species. | ||||||||
| partial veil | absent. | ||||||||
| lamella edge tissue | sterile. | ||||||||
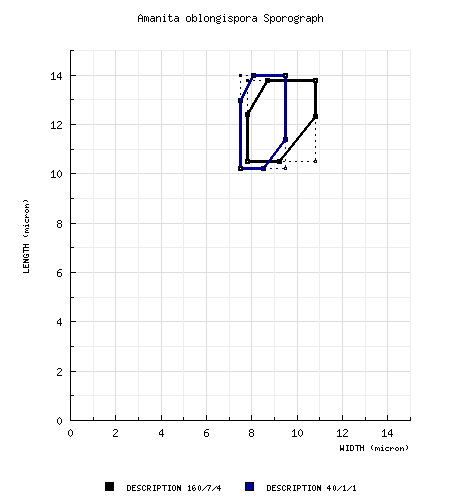
| basidiospores |
from protolog: [160/7/4] (8.2-) 10.5 - 13.8 (-16.2) × (7.2-) 7.8 - 10.8 (-13.5) µm, (L = 11.5 - 12.6 µm; L' = 12.1 μm; W = 8.5 - 10.1 µm; W' = 9.2 μm; Q= (1.07-) 1.14 - 1.59 (-2.16); Q = 1.19 - 1.45; Q' = 1.33), hyaline, colorless, thin-walled, smooth, inamyloid, broadly ellipsoid to ellipsoid, occasionally elongate, infrequently cylindric, occasionally subventricose to ovoid, often adaxially flattened; apiculus sublateral, cylindric, broad at times; contents granular to mono- to multiguttulate; white in desposit. from holotype only (RET): [40/1/1] (8.2-) 10.2 - 14.0 (-16.2) × (7.2-) 7.5 - 9.5 (-10.0) µm, (L = 12.2 µm; W = 8.5 µm; Q= (1.14-) 1.20 - 1.73 (-2.16); Q = 1.45). [Note: Spore measurements for the holotype are segregated because I have come to think that one or more of the paratypes may have to be excluded from the species.—RET] | ||||||||
| ecology | from protolog: Infrequent. Under mixed broad-leaved trees including Quercus spp. in humicolous acid soil. | ||||||||
| material examined | from protolog: ITALY: SARDINIA—Prov. Cagliari - "la Cannoneris" St. For., Punta Sebera, 25.x.1986 M. Contu 86 10 25/05 (holotype, CAG). Prov. Nuoro - Laconi, 20.vi.1992 M. Contu 92/37 (paratype, CAG), 92/38 (paratype, CAG; paratype, RET, fragments), 92/43 (paratype, CAG). | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.