


| name | Amanita marmorata |
| name status | nomen acceptum |
| author | Cleland & E.-J. Gilbert |
| english name | "Marbled Death Cap" |
| synonyms |
=Amanita reidii Eicker & Greuning
=Amanita marmorata subsp. myrtacearum O. K. Mill., Hemmes & G. Wong |
| images |
 1. Amanita marmorata, Blue Mountains, New South Wales, Australia. RET 623-7  2. Amanita marmorata, Blue Mountains, New South Wales, Australia. RET 623-7  3. Amanita marmorata, Oahu, Hawaii, U.S.A. RET 044-6  4. Amanita marmorata, Oahu, Hawaii, U.S.A. RET 044-6  5. Amanita marmorata, Oahu, Hawaii, U.S.A. RET 044-6 |
| intro | This description of Amanita marmorata is based on the original descriptions of the three synonyms above, which appear in the following works: Gilbert (1941), Eicker et al. (1993), and O. K. Miller et al. (1996). |
| cap | The cap of Amanita marmorata is 30 - 72 (-95) mm wide, with a densely flecked (not quite virgate or streaked) surface having a mixture of small areas colored brown or gray or gray-brown, (sometimes slate gray or dark gray-brown in the center, sometimes with a yellowish tint in the center, sometimes with all pigmented flecks gray in age) and white or whitish, creating a marbled appearance, shallowly campanulate to shallowly convex to planar, with a nonstriate, nonappendiculate margin. The universal veil is almost always absent. If present, it forms a white, membranous patch. |
| gills | The gills are crowded, free to narrowly attached, white to creamy white, and rather narrow. |
| stem | The stem is 30 - 70 × 5 - 15 mm, silky, white to dirty white, farinose above the ring, sometimes with zig-zag ("flame" or "zebroid") markings below the ring. The ring is apical, skirt-like, white, and membranous. The stem bears an ample, weakly membranous, limbate volva arising from near the top of the stem's bulbous base (which is up to 22 mm wide). |
| odor/taste | This mushroom is said to have a "strong" odor. It is also contains amatoxins and is deadly POISONOUS. |
| spores | The spores measure (6.8-) 7.5 - 10.0 (-11.8) × (5.8-) 6.2 - 8.0 (-9.5) µm, are subglobose to broadly ellipsoid to ellipsoid (rarely elongate) and are amyloid. Clamps are not found at bases of the basidia. |
| discussion |
Amanita marmorata is deadly POISONOUS. The species contains amatoxins. Amanita marmorata was originally described from the state of South Australia. It has been exported with Australian plants to both South Africa and the Hawaiian Islands, USA. In each of these localities it received a new name. For similar taxa, see the page for A. phalloides (Fr. : Fr.) Link. Plants reportedly associated with the present species in Hawaii include Eucalyptus, Casuarina, Araucaria, and Melaleuca (Miller et. al., 1996). In South Africa, Eucalyptus is reported as the sole associated genus (Eicker et al., 1993). The material that I have examined (for example, for spore measurements above) is largely from Hawaii. The editors of these pages are both interested in obtaining well-documented, well-dried collections from Australia and South Africa as well as Hawaii.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita marmorata | ||||||||||||||||||||||||
| author | Cleland & E.-J. Gilbert in E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl. (2): 329, tab. 45. | ||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||
| english name | "Marbled Death Cap" | ||||||||||||||||||||||||
| synonyms |
≡Amanita [?Amanitina] marmorata Cleland & E.-J. Gilbert nom. nud. in E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 78, 94, 96, tab. 33 (figs. 5-6), tab. 34 (figs. 1-3). [Lacking Latin diagnosis, specification of holotype, etc. ICBN §36.1, §37.1, etc.]
=Amanita reidii Eicker & Greuning in Eicker, Greuning & D. A. Reid. 1993. Mycotaxon 47: 443, figs. 1-8.
=Amanita marmorata subsp. myrtacearum O. K. Mill., Hemmes & G. Wong. 1996. Mycologia 88: 140, figs. 1-7. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||||||
| MycoBank nos. | 284063, 360162, 414508 | ||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||
| holotypes | A. reidii—PRUM. A. marmorata subsp. myrtacearum—VPI. | ||||||||||||||||||||||||
| lectotypes | A. marmorata—AD. | ||||||||||||||||||||||||
| lectotypifications | A. marmorata—Reid. 1980. Austral. J. Bot., Suppl. Ser. 8: 39. | ||||||||||||||||||||||||
| type studies | A. marmorata—Reid. 1980. Austral. J. Bot., Suppl. Ser. 8: 38, fig. 24. | ||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon or of one of its synonyms and not cited as the work of another researcher is based upon original research by R. E. Tulloss. | ||||||||||||||||||||||||
| pileus |
from protolog: 35 mm wide, white at first, becoming radially marbled with grayish brown, globose then convex, slightly viscid when moist; context not described; margin nonstriate; universal veil not described. from Reid (1980): universal veil as inconspicuous fragments on margin of lectotype, thin, "felty". from Eicker et al. (1993) [A. reidii]: 30 - 72 mm wide, "blotchily" gray-brown, sometimes darker gray-brown to blackish slate-gray over disc, sometimes paler toward margin, often developing irregularly distributed "pallid dirty white blotches," "innately radially streaky" (most obvious toward margin), shallowly campanulate to convex, becoming shallowly convex or planar, never observed to be viscid (per discussion); context not described; margin not described; universal veil completely lacking. from Miller et al. (1996): (35-) 42 - 55 (-95) mm wide, white at first (sometimes with a yellowish hue over disc), soon becoming marbled light gray to gray (5C3 to Light Gray to Smoke Gray) to occasionally brownish gray (5C3 to Light Drab to Drab) over entire surface or not quite extending to margin, sometimes entirely gray with age, convex to broadly convex, planar in age; context firm and white; margin not described; universal veil not described. | ||||||||||||||||||||||||
| lamellae |
from protolog: attachment not described, close, white, at times weakly tinted pink; lamellulae not described. from Eicker et al. (1993) [A. reidii]: adnexed to almost free; density not described; white to creamy white. from Miller et al. (1996): narrowly free, close, white; lamellulae evenly distributed, one between each pair of otherwise adjacent lamellae, of diverse lengths [per photo (Miller et al. 1996: fig. 1)], plentiful [per same figure]. [Note: Immediately above the text stating that lamellulae appear in a single "tier" is a photograph clearly showing that the lamellulae vary in length.—ed.] | ||||||||||||||||||||||||
| stipe |
from protolog: white, sometimes silky, powdery above; bulb present; context becoming hollow; partial veil superior, membranous, pendent, ample; universal veil as limbate volva, membranous, ample, flaccid. from Eicker et al. (1993) [A. reidii]: 30 - 70 × 0.5 - 1.5 mm (width measured at apex, [length apparently including that of bulb—ed.]), white to dirty white, with "fibrillose scaly zig-zag markings" below partial veil; bulb up to 22 mm wide, clavate ("nonbulbous"); context not described; partial veil apical, pendent, white, membranous; universal veil as "saccate" volva, "although free from .. stipe sheathing it rather closely." from Miller et al. (1996): 40 - 80 (-100) × (3-) 5 - 10 (-15) mm, white, cylindric or narrowing upward, dry, covered with slightly raised loose fibrils; bulb as clavate base; context white, soft in center, firm otherwise. RET: 35 - 90 × 13 - 16 mm, white, narrowing upward, finely longitudinally striatulate; bulb 24 - 43 × 25 - 38, subglobose to clavate; partial veil superior, white, membranous, pendent, skirt-like, persistent; universal veil as limbate volva, white, membranous, persistent, with upper tip of limb up to 57 mm from bottom of bulb. | ||||||||||||||||||||||||
| odor/taste |
from protolog: Odor strong. Taste not recorded. from Eicker et al. (1993) [A. reidii]: not described. from Miller et al. (1996): Odor mild, fungoid. Taste not recorded. | ||||||||||||||||||||||||
| macrochemical tests |
from Miller et al. (1996): 10% KOH aq. soln. - bright yellow on pileipellis and along the interface between lamella and pileus context. 3% KOH aq. soln. - paler reaction than above. | ||||||||||||||||||||||||
| pileipellis |
from Eicker et al. (1993) [A. reidii]: not described. from Miller et al. (1996): more tightly interwoven than pileus context; filamentous hyphae 3 - 7 μm wide, cylindric to "slightly swollen"; clamps lacking. RET: 75 - 110 μm thick, in mass hyaline to pale yellowish, slightly gelatinized only at very surface; filamentous undifferentiated hyphae 2.0 - 11.2 μm wide, branching, with walls up to 0.5± μm thick, in scalp view many subradially arranged and densely interwoven and some criss-crossed and loosely interwoven; vascular hyphae 7.6 - 8.0 μm wide, sinuous, uncommon. | ||||||||||||||||||||||||
| pileus context |
from Eicker et al. (1993) [A. reidii]: not described. from Miller et al. (1996): filamentous hyphae (2.5-) 4.0 - 11.0 (-15.0) μm wide, cylindric to "slightly swollen"; acrophysalides not described; vascular hyphae with yellow contents, forming "sparse network; clamps lacking. RET: filamentous undifferentiated hyphae 2.5 - 18.0 μm wide, branching, singly or in fascicles, loosely interwoven, plentiful, thin-walled or with wall slightly thickened (up to 0.5± μm thick), with occasionl intercalary partially inflated segments up to 17.5 μm wide; acrophysalides plentiful, clavate to narrowly clavate to ellipsoid (e.g., 67 - 80 × 25 - 47 μm or possibly larger), with walls thin or slightly thickened (up to 0.5± μm thick) or rarely up to 1.0 μm thick; vascular hyphae not observed. | ||||||||||||||||||||||||
| lamella trama |
from Eicker et al. (1993) [A. reidii]: not described. from Miller et al. (1996): divergent; filamentous hyphae 6 - 18 μm wide; inflated cells not described; vascular hyphae scattered, with yellow contents; clamps lacking. RET: bilateral, divergent, with distinct central stratum; wcs = 40 - 55 μm (moderate to good rehydration); filamentous undifferentiated hyphae 2.2 - 7.5 μm wide; inflated cell at times bridging entire distance between central stratum and subhymenium, thin-walled, ovoid to irregular, up to 32 × 13.0 μm, terminal divergent inflated cells not observed; vascular hyphae not observed; clamps not observed. | ||||||||||||||||||||||||
| subhymenium |
from Eicker et al. (1993) [A. reidii]: with basidia arising from "rounded" or "wedge-shaped" cells. from Miller et al. (1996): several cells deep; cells "irregular," 9 - 14 μm "diam"; clamps lacking. [Note: It is not clear what the "diameter" of an "irregular" cell might be. The authors omit the word "inflated", making interpretation of the form of the cells of the subhymenium difficult.—ed.] RET: pseudoparenchymatous (cellular), with cells in 2 - 3 (-4) layers; wst-near = 15 - 25 μm; wst-far = 25 - 35 μm; with cells predominantly inflated, usually with greatest dimension < 9.0 μm, sometimes up to 14.0 μm, clavate to ellipsoid to subglobose to slightly irregular, with basidia arising from cells of all forms as well as from very short partially inflated hyphal segments. | ||||||||||||||||||||||||
| basidia |
from Reid (1980): up to 38 × 11.6 μm; clamps not observed. from Eicker et al. (1993) [A. reidii]: 36 - 50 × 7 - 11 μm, 4-sterigmate; clamps lacking. from Miller et al. (1996): 32 - 43 × 8 - 11 μm, 4-sterigmate, with sterigmata 3 - 4 μm long; clamps absent. RET: 24 - 42 × 7.8 - 12.0 μm, 4-sterigmate; clamps not observed. | ||||||||||||||||||||||||
| universal veil |
from Eicker et al. (1993) [A. reidii]: On pileus: absent. On stipe base, apparently interior layer: filamentous hyphae up to 4 μm wide, with wall thin "but distinct," with intercalary often fusiform inflated segments up to 200 × 35 μm; inflated terminal cells absent ("devoid of spherocysts"); vascular hyphae occasional in some collections; clamps lacking. from Miller et al. (1996): filamentous hyphae 3 - 15 (-20) μm, infrequently physalomitic [i.e., having inflated segments]; inflated cells 50 - 100 × 14 - 17 μm, scattered, terminal, similar to those in the partial veil. [Note: Since Miller distinguishes inflated cells that are terminal on hyphae from "physalomitic" hyphae, we can assume that he is referring to intercalary inflated segments in using the term "physalomitic."—ed.] RET: On pileus: absent. On stipe base, exterior surface layer: gelatinized just at surface; longitudinally oriented filamentous undifferentiated hyphae 1.2 - 11.8 μm wide, densely interwoven, branching, with walls thin or slightly thickened (up to 0.5± μm thick), branching, with scattered intercalary partially inflated cells (narrowly clavate to subfusiform, up to 100 × 24 μm) with slightly thickened walls; inflated cells ??; vascular hyphae not observed. On stipe base, interior layer: filamentous undifferentiated hyphae 3.2 - 12.0 (-21±?) wide, dominant, branching, often in fascicles, moderately densely interwoven, thin-walled or frequently with walls slightly thickened (< 0.5 μm thick), with scattered intercalary cells up to 125 × 26 μm with walls thin or up to 1.0+ μm thick; inflated cells scattered, sometimes in small clusters, terminal singly,thin-walled, ellipsoid to narrowly clavate to broadly clavate (up to 93 × 36 μm or larger) or subglobose (e.g., 63 × 54 μm) or cylindric (e.g., 55 × 27 μm); vascular hyphae not observed; clamps not observed. On stipe base, inner surface layer: as thin gelatinized layer similar to interior. | ||||||||||||||||||||||||
| stipe context |
from Eicker et al. (1993) [A. reidii]: not described. from Miller et al. (1996): not described. RET: longitudinally acrophysalidic; filamentous undifferentiated hyphae 3.5 - 10.5 μm wide, plentiful, branching, thin-walled; acrophysalides very slender, dominant, thin-walled or with walls slightly thickened (up to 0.5± μm thick), up to 309 × 34 μm; vascular hyphae not observed; clamps not observed. | ||||||||||||||||||||||||
| partial veil |
from Eicker et al. (1993) [A. reidii]: "hyphal." from Miller et al. (1996): filamentous hyphae 3 - 10 μm wide, interwoven, often branched; inflated cells numerous, narrowly clavate, 30 - 60 × 8 - 11 μm, terminal singly; vascular hyphae not described; clamps absent. RET: filamentous undifferentiated hyphae 2.0 - 8.5 μm wide, dominating, frequently branching, with some branches coiled or tangled, interwoven, often in fascicles with subradial orientation, occasionally slightly gelatinized near surfaces, with terminal segments very slightly expanded at tips; inflated cells common, thin-walled, barely inflated (intergrading with barely expanded tip segments), terminal singly, up to 37 × 16.0 μm; vascular hyphae not observed. | ||||||||||||||||||||||||
| lamella edge tissue |
from Eicker et al. (1993) [A. reidii]: inflated cells 20 - 36 × 10 - 16 μm, hyaline, thin-walled, clavate or ovate; "In the Bergvliet material...[as] thick layer of dissociating chains of thin-walled, hyaline, globose or ovate elements up to 30μm...[×]... 23 μm...." [Note: It is curious that the authors could describe the tissue of the Bergvliet material in the above detail and still misuse the term "cheilocystidia" for isolated elements on gill edges of other specimens.—ed.] from Miller et al. (1996): as a sterile layer over [edge of] lamellae; inflated cells 20 - 44 × 10 - 14 (-38) μm, broadly clavate. [Note: The cells are called "cheilocystidia" again in this case. The comment immedately above applies here again. By definition, cystidia do not form continuous layers.—ed.] | ||||||||||||||||||||||||
| basidiospores |
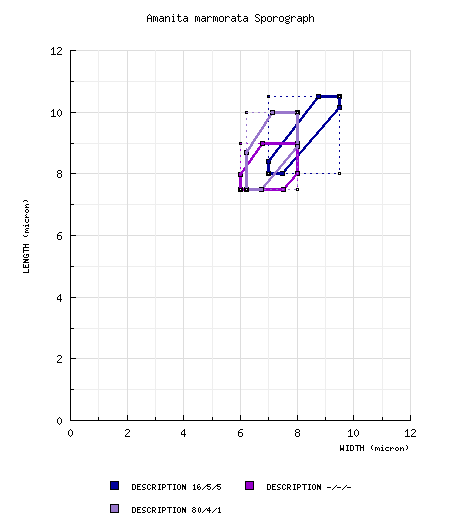
from protolog [tab. xxxiii (fig. 5-6) and tab. xxxiv (figs. 1-3) of Gilbert (1940)]: [16/5/5] (7.5-) 8.0 - 10.5 (-11.2) × (6.8-) 7.0 - 9.5 (-10.0) μm, (L' = 9.3 μm; W' = 8.2 μm; Q = (1.05-) 1.07 - 1.20 (-1.22); Q' = 1.13). [Note: The scale drawings of spores in Gilbert (1940) have proven to be a very reliable source of spore measurements for many taxa. Data is retained from these spores only in the cases in which a spore is shown in strictly lateral view.—ed.] from Reid (1980): From spore print: 7.0 - 9.0 × 6.0 - 7.5 μm; color in deposit not recorded. From gill: 7.0 - 9.5 × 5.0 - 7.0 μm. [Note: As is common in the work of Reid (see A. griseovelata), the spore measurements and shapes are not treated as values in continua. This results in spore data for which Q values are difficult to estimate and for which sporographs are, consequently, not necessarily of proper form. No sporograph is generated.] from Eicker et al. (1993) [A. reidii]: From spore print: (7.0-) 8.0 - 9.5 (-10) × (6.0-) 6.2 - 9.0 μm; color in deposit not recorded. [Note: As is common in the work of Reid (see A. griseovelata), the spore measurements and shapes are not treated as values in continua. This results in spore data for which Q values are difficult to estimate and for which sporographs are, consequently, not necessarily of proper form. In this case, we have added to the single set of measurements for spores from a print, the extremes values from the two additional sets of measurements reported based on spores found on lamellae. No sporograph is generated.—ed.] from Miller et al. (1996) [A. marmorata subsp. myrtacearum]: [-/-/-] (6.7-) 7.5 - 9.0 (-12.0) × (5.5-) 6.0 - 8.0 (-10.5) μm, (Q = 1.0 - 1.33; Q' = 1.15), hyaline, thin-walled, amyloid, globose to subglobose to broadly ellipsoid to ellipsoid; apiculus inamyloid; contents not described; white in deposit. [Note: The apiculus is always inamyloid in Amanita independent of the remainder of the spore to Melzer's reagent. Because none of the 10 spores that are illustrated are drawn in lateral view, there is reason to believe that the spore data may have been collected from spores that were not all in lateral view when measured. This would cause the measured lengths of spores to be depressed, in turn, causing the average value of Q computed from these measurements to be depressed—spores would appear more nearly globose on average than they would appear if measurements were only made on spores in lateral view—the standard method supported by editors. The two slanted sides of the sporograph would be shifted downward toward the line at a 45° angle to the horizontal axis, which represents a Q value = 1.0. The reader should compare the sporograph generated from the Miller et al. data with the sporograph generated from the data below.—ed.] RET: [80/4/1] (6.8-) 7.5 - 10.0 (-11.8) × (5.8-) 6.2 - 8.0 (-9.5) μm, (L = 8.1 - 9.1 μm; L’ = 8.6 μm; W = 6.8 - 7.2 μm; W’ = 7.0 μm; Q = (1.06-) 1.11 - 1.40 (-1.67); Q = 1.17 - 1.29; Q’ = 1.22), hyaline, colorless, ?, smooth, amyloid, broadly ellipsoid to ellipsoid, occasionally subglobose, adaxially flattened, with occasional "giant" spores on presumably young specimens; apiculus sublateral, ?; contents ?; color in deposit not recorded. | ||||||||||||||||||||||||
| ecology |
from protolog: Terrestrial. from Eicker et al. (1993) [A. reidii]: Under [alien] Eucalyptus claeziana or under inidentified[, alien] Eucalyptus sp. [Note: The authors raise the possibility that the species might have been imported to South Africa from Australasia.—ed.] from Miller et al. (1996) [A. marmorata subsp. myrtacearum]: In association with the following [alien] trees: Eucalyptus robusta, E. saligna. E. sp., Casuarina equisetifolia (where it may occur in a pure stand), Araucaria columnaris, and Melaleuca quinquenervia. It is associated with other Australian species of the Myrtaceae in the National Tropical Botanical Garden, Lawai, Kauai. Since at least some species of Araucaria are known to be endomycorrhizal, A. marmorata may not be mycorrhizal with trees in that genus. All putative hosts in Hawaii were imported from eastern states of Australia. Other Hawaiian site: Under Araucaria heterophylla (Norfolk Island Pine) and Casuarina. | ||||||||||||||||||||||||
| material examined |
from Reid (1980): AUSTRALIA: NEW SOUTH WALES—Sydney, Neutral Bay, 5.iv.1914 J. B. Cleland s.n. (lectotype, ADW 9252 => AD). [Note: In the protolog, a type collection is not specified (per Reid), and the state in which Sydney is located is incorrectly stated to be South Australia.] from Eicker et al. (1993) [A. reidii]: SOUTH AFRICA: GAUTENG—ca. Pretoria, Melkrivier, Hideaway Game Farm, 29.iv.1990 V. van Greuning & D. A. Reid s.n. (holotype, PRUM 3179). MPUMALANGA—Belfast, Pineglades, 8.ii.1991 unkn. coll. s.n. (paratype, herb. unkn.); Sabie, Bergvliet St. For., 7.xii.1984 G. C. A. van der Westhuizen s.n. (paratype, PREM 48618). from Miller et al. (1996) [A. marmorata subsp. myrtacearum]: U.S.A.: HAWAII—Hawaii - Hilo, Univ. of Hawaii at Hilo, 18.i.1990 D. E. Hemmes 207 (paratype, BISH), D. E. Hemmes 484 (paratype, BISH), 25.vii.1990 O. K. Miller 24545 (holotype, VPI), 17.ix.1990 D. E. Hemmes 124 (paratype, VPI); Paradise Pk. [19°34'27" N/ 154°57'30" W], 1.viii.1990 D. E. Hemmes & O. K. Miller 24560 (paratype, VPI); MacKenzie St. Pk. [19°26'24" N/ 154°52'10" W], 10.xii.1991 D. E. Hemmes 226 (paratype, BISH). Kauai - Lawai, Nat. Tropical Botanical Gardens [21°54'29" N/ 154°30'43" W, 129 m], 23.xi.1990 D. E. Hemmes & G. J. Wong [Wong 871] (paratype, VPI), 20.xii.1990 D. E. Hemmes 231 (paratype, BISH); Kokee, 3.x.1994 D. E. Hemmes 597 (VPI), 4.x.1994 D. E. Hemmes 616 (paratype, VPI). Lanai - Lanaihale Tr., 15.i.1985 G. J. Wong 800 (paratype, BISH). Maui - Kaumahina St. Wayside, 14.vi.1992 D E. Hemmes 218 (paratype, BISH); Maluhia Boy Scout Camp, 24.vii.1991 D. E. Hemmes 209 (paratype, BISH); Waihee Ridge Tr., 26.vii.1991 D. E. Hemmes 210 (paratype, BISH). Molokai - along Molokai For. Reserve Rd., 25.iii.1991 G. J. Wong 1024 (paratype, BISH); Palaau St. Pk. [21°10'25" N/ 157°00'30" W], 2.xii.1991 G. J. Wong 800 (paratype, BISH). Oahu - Aiea, Aiea Loop Tr., 30.xi.1990 G. J. Wong 906 (paratype, BISH); Honolulu, Manoa Cliffs Tr., 4.x.1990 G. J. Wong 860 (paratype, BISH), 862 (paratype, BISH); Honolulu, Manoa Valley, 4.xi.1984 D. J. Friend s.n. (paratype, BISH); Honolulu, Nuuanu Pali St. Wayside [21°21'59" N/ 157°47'38" W, 361 m], 16.i.1992 D. E. Hemmes 214 (paratype, BISH); Honolulu, Waahila Ridge St. Recreation Area [21°18'25" N/ 157°47'50" W], 22.iv.1989 G. Wong & O. K. Miller [Miller 23553] (paratype, VPI); Waianae, Peacock Flats Campgrd., 28.xii.1990 G. J. Wong 975 (paratype, BISH); Waianae, Kaena Pt. Satellite Tracking Stn. [ca. 21°33'49" N/ 158°14'20" W], 14.xii.1991 G. J. Wong 1146 (paratype, BISH). from Wood (1997): AUSTRALIA: NEW SOUTH WALES—Sydney - Oatley, 11.iv.1994 A. E. & J. E. Wood s.n. (UNSW 94/4). Unkn. LGA - Mooball St. For., SE of Murwillumbah, Mooball St. For. [28°23'00" S/ 153°28'00" E], 17.iii.1983 J. J. Bruhl s.n. (UNSW 83/217); Cessnock, 4.iv.1983 A. Chalk s.n. (USNW 83/346B). RET: AUSTRALIA: NEW SOUTH WALES— City of Lithgow, Blue Mountains, Little Hartley [33°33'35.63" S/ 150°12'07.99" E, 819 m], 12.iv.2014 Lucy Albertella s.n. [mushroomobserver #163271] (RET 623-7, nrITS & nrLSU seq'd.). SOUTH AFRICA: GAUTENG PROV.—Muldersdrift, Avianto [26.0236° S/ 27.8416° E, 1433 m], 11.iv.2015 Liz Popich s.n. [mushroomobserver #202637] (RET 685-9, rnITS & nrLSU seq'd.). U.S.A.: HAWAII—Oahu - above Manoa Valley, St. Louis Heights, | ||||||||||||||||||||||||
| discussion |
At the time of the descriptions of A. reidii and the putative subspecies of Miller et al., there were no known collections of this species from Australia of more recent data than Cleland's original collections. Wood (1997) published on four collections of which at least three may belong to the present taxon. Minimal microscopic information is provided; and, as noted elsewhere on this site, Wood's use of Q in the 1997 publication was idiosyncratic and cannot support production of a sporograph. We have included the three collections that he does not himself question in the material examined data field, above. In as yet unpublished, molecular research Dr. H. Hallen (then of Michigan St. Univ.) found that the present species contains amanitin and, among those taxa of sect. Phalloideae that she included in her research, the present species was basal to the clade including all sequenced taxa known to contain one or more amanitins. This suggested to her the hypothesis that the amanitins evolved in a single mutation event in Australia or a larger fragment of Gondwana that existed at the time. The following figure provides comparison of the sporographs of the present and A. phalloides to which A. marmorata has been compared by several authors: It is clear that distinguishing between the two taxa on the basis of spore measurements alone would not be satisfactory. Instead Eicker et al. focused attention on (1) the clear difference in cap pigmentation, range of size of the basidiomes in the two taxa, (2) differences in bulb shape between the two taxa (a clavate form for the bulb would be very unusual in A. phalloides), (3) difference in known sets of mycorrhizal symbionts of the taxa, and (4) what the editors interpret as a statement the free limb of the volva in the present species is longer as a proportion of the distance from the tip of that limb to the base of the bulb than is the case in A. phalloides. There are additional anatomical characters that distinguish the two taxa. Reid (1980) had previously noted that the walls of the hyphae in the interior of the universal veil of A. phalloides are often thickened and had illustrated this phenomenon (Reid 1980: fig. 59). In fact, RET confirms that these hyphae in A. phalloides are dominantly thick-walled and the wall thickness can exceed twice the thickness RET observed in the thickest hyphal walls in the universal veil interior of the present taxon. Since Eicker et al. mention the inflated intercalary segments of hyphae in A. reidii as a possible character distinguishing their new taxon from A. alauda (see below), it is worth noting that these intercalary cells are also reported on this site by RET in the universal veil of A. phalloides. The following figure provides comparison of the sporographs of the present species and A. alauda (described from tropical "jungle" in Singapore) to which A. marmorata has been compared by Eicker et al. in the protolog of A. reidii: The spores of A. alauda are shorter than those of the present species while the range of spore widths is very similar in the two taxa. Consequently, the shape of the spores of A. alauda is notably more globose than is the case in A. marmorata, and spore measurement data is one of the characters useful in segregating the present species from A. alauda. Among other characters segregating the little known A. alauda from the present species Eicker et al. list the following: (1) darker pigment nearer to the pileus margin, (2) a central umbo in a central depression on the pileus, (3) viscidity of the pileus, (4) the supposed (but unconfirmed) absence of fusiform intercalary elements of the interior of the universal veil, and (5) the difference in known habitat ecology. The observed lack of visicidity of the cap in the original material of A. reidii is consistent with RET's observation that the pileipellis in the Dale collectons from Oahu were barely gelatinized at the surface. | ||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita marmorata |
| name status | nomen acceptum |
| author | Cleland & E.-J. Gilbert |
| english name | "Marbled Death Cap" |
| images |
1. Amanita marmorata, Blue Mountains, New South Wales, Australia. RET 623-7 2. Amanita marmorata, Blue Mountains, New South Wales, Australia. RET 623-7 3. Amanita marmorata, Oahu, Hawaii, U.S.A. RET 044-6 4. Amanita marmorata, Oahu, Hawaii, U.S.A. RET 044-6 5. Amanita marmorata, Oahu, Hawaii, U.S.A. RET 044-6 |
| photo |
Lucy Albertella - (1-2) Blue Mountains, New South Wales, Australia. (RET 623-7) [Note: These images are used with permission. They may be view untrimmed and in original size here.] Jon Dale - (3-5) Oahu, Hawaii, U.S.A. (RET 044-6) Liz Popich - (6-7) Avianto, Muldersdrift, Gauteng, South Africa (RET 685-9) [Note: Button. Untrimmed and unedited images may be found at mushroomobserver.org/202637] |
| name | Amanita marmorata |
| bottom links |
[ Keys & Checklists ] |
| name | Amanita marmorata |
| bottom links |
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.