


| name | Amanita lesueurii | ||||||||||||
| author | E. M. Davison in Davison et al. 2013. Nuytsia 23: 591, figs. 1-2. | ||||||||||||
| name status | nomen acceptum | ||||||||||||
| english name | Lesueur's Lepidella | ||||||||||||
| etymology |
Latin genitive of the name Lesueur. protolog: "The epithet celebrates Charles-Alexandre Lesueur (1778–1846) botanical artist and natural history painter on the Baudin expedition to Australia (1800–1804)." | ||||||||||||
| MycoBank nos. | 800268 | ||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||
| holotypes | PERTH 08351325 | ||||||||||||
| intro |
The field may contain magenta text presenting data from a type study
and/or revision of other original material cited in the protolog of the present taxon.
Macroscopic descriptions in magenta are a combination of data from the protolog and
additional observations made on the exiccata during revision of the cited original
material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is based directly from the protolog of the present taxon. protlog: Basidiomes small to medium-sized. | ||||||||||||
| pileus | protolog: 30 - 72 mm wide, up to 10 mm thick, initially white becoming clay-buff to vinaceous-buff (6B2–C3) with age, without surface staining or bruising, initially convex becoming plane, slightly tacky when moist; context white bruising pale vinaceous-buff (6B2); margin non-striate, appendiculate, becoming decurved with age; universal veil as adnate, aerolate, floccose-felted warts or patches becoming adpressed with age, white at first, aging pale vinaceous-buff to grey (6B–C2). | ||||||||||||
| lamellae | protolog: adnate to adnexed, close to subdistant, white, unchanging, 5 - 10 mm broad, with margin concolorous and occasionally fimbriate; lamellulae truncate (shortest), attenuate (longest), scarce to plentiful, in several lengths. | ||||||||||||
| stipe | protolog: 16 - 30 × 8 - 21 mm, cylindric or narrowing upwards, solid, white, slightly floccose below partial veil; bulb 5–20 × 10–22 mm, obconic or tapered or turbinate, encrusted with soil; context as in pileus; partial veil inferior to median to superior, descendant, membranous, adpressed, white unchanging, persistent or disappearing with age; universal veil not apparent or white and as slight fragile membranous limb or small marginate rim on bulb. | ||||||||||||
| odor/taste | protolog: Odor none when young, strong when old. Taste not recorded. | ||||||||||||
| macrochemical tests |
none recorded. | ||||||||||||
| pileipellis | protolog: difficult to delimit in young specimens, up to 250 μm thick in older specimens, initially colourless, becoming pale brown in NH4OH; filamentous hyphae 3 - 10 μm wide with thick gelatinizing walls, radially orientated, periclinal, occasionally interwoven; inflated cells not observed; vascular hyphae 5–10 μm wide, pale yellow, infrequent, occasionally sinuous; clamps not observed. | ||||||||||||
| pileus context | protolog: filamentous, hyphae 3 - 30 μm wide, with widest constricted at septa, thin-walled, hyaline, dominant; acrophysalides up to 220 × 20 μm, thin-walled, cylindric or clavate, colorless; vascular hyphae 3 - 10 μm wide, occasionally branching, pale yellow, very infrequent to locally frequent, occasionally sinuous; clamps not observed. | ||||||||||||
| lamella trama | protolog: bilateral, divergent. Central stratum (when well rehydrated) comprising 12 - 18% of distance between bases of basidia on opposing hymenial surfaces; undifferentiated hyphae 2 - 18 μm wide, with widest constricted at septa, thin-walled, hyaline; inflated cells not observed; vascular hyphae 2 - 6 μm wide, rare, pale yellow, occasionally sinuous; clamps not observed. Subhymenial base with initial angle of divergence from 15° to 30° from central stratum; filamentous hyphae following smooth broad curve to subhymenium; filamentous hyphae 4 - 25 μm wide, frequently branched, thin-walled, hyaline; inflated cells infrequent, colorless, up to 80 × 30 μm, clavate or fusiform or ovoid; vascular hyphae 3 - 5 μm wide, rare, pale yellow, occasionally sinuous; clamps not observed. | ||||||||||||
| subhymenium | protolog: with basidia arising terminally from barely inflated to pyriform hyphal segments 5 - 12 μm wide; clamps not observed. [Note: Per illustration, Bas' terms "ramose" or "subinflated ramose" would appear to apply to the subhymenium.—ed.] | ||||||||||||
| basidia | protolog: [100/5/5] (42-) 44 - 63 (-75) × (9-) 10 - 13 (-14) μm, thin-walled, colorless, about 86% 4-spored, about 6% 3-spored, about 4% 2-spored, about 3% 1-spored, with sterigmata up to 20 × 3 μm wide, with most less than 6 × 2 μm, clamps not observed. | ||||||||||||
| universal veil | protolog: On pileus: not layered, with elements somewhat erect or irregularly disposed; filamentous hyphae 3 - 10 μm wide, colorless or pale brown; inflated cells dominant, ellipsoid or ovoid or pyriform (up to 115 × 60 μm) in basal part, ellipsoid or globose (up to 45 × 40 μm) in upper part, terminal singly or in chains of up to 3 cells, with walls thin or slightly thickened and gelatinizing, contents colourless or light brown or darker in collapsed cells; vascular hyphae 3 - 10 μm wide, frequently branching, sinuous, colorless or pale yellow or brown, most frequent nearest pileipellis; clamps not observed. On stipe base: elements without clear orientation, pale yellow or pale brown in NH4OH; filamentous hyphae 3 - 12 μm wide hyaline, gelatinizing; inflated cells plentiful to dominant, ellipsoid or globose or ovoid or pyriform or clavate or fusiform (up to 65 × 55 μm) in terminal chains of up to 4 cells, with walls thin or slightly thickened; vascular hyphae 3 - 5 μm wide, pale yellow, infrequent; clamps not observed. | ||||||||||||
| stipe context | protolog: longitudinally acrophysalidic; filamentous hyphae 3 - 15 μm wide, hyaline; acrophysalides dominant, up to 200 × 40 μm, mainly clavate, thin-walled, colorless; vascular hyphae 5 - 10 μm wide, occasionally branching, colourless or pale yellow, not particularly concentrated at stipe apex; clamps not observed. | ||||||||||||
| partial veil | protolog: filamentous hyphae 3 - 12 μm wide, hyaline, radially orientated, dominant, disarticulating at septa; inflated cells infrequent, clavate, up to 25 × 10 μm, thin-walled, colorless; vascular hyphae 2 - 8 μm wide, pale yellow, frequent; clamps not observed. | ||||||||||||
| lamella edge tissue | protolog: sterile; inflated cells pyriform or clavate or capitate, 18 - 50 × 10 - 20 μm, colorless, infrequent to frequent; clamps not observed. | ||||||||||||
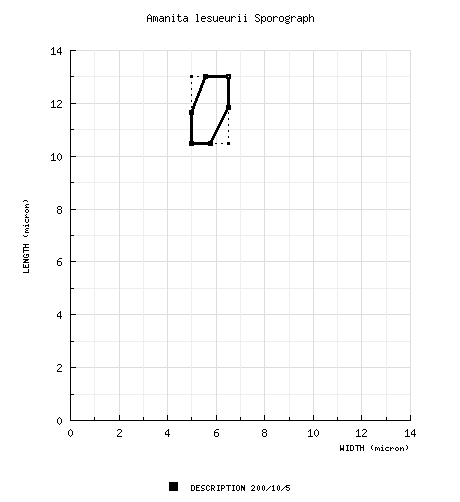
| basidiospores | protolog: [200/10/5] (9.0-) 10.5 - 13.0 (-14.0) × 5.0 - 6.5 (-7.0) μm, (L = 11.1 - 12.4 μm; L’ = 11.7 μm, W = 5.5 - 6.2 μm; W’ = 5.8 μm; Q = (1.62-) 1.82 - 2.33 (-2.60); Q = 1.90 - 2.26; Q’ = 2.03), hyaline, colorless, with walls slightly thickened, smooth, amyloid, elongate to cylindric; apiculus sublateral, short, cylindric to truncate-conic, up to 1 × 2 μm, truncate; contents monoguttulate; white in deposit. | ||||||||||||
| ecology | protolog: Solitary to gregarious. In sandy soil in kwongan vegetation and low mulga woodland over scrub; nearby plants include Allocasuarina humilis, Corymbia calophylla, Acacia aneura, Daviesia sp., Eremophila sp. and Melaleuca sp. Occurs in the Geraldton Sandplains and Yalgoo bioregions (Department of the Environment [Australia] 2013). Fruiting June to September. | ||||||||||||
| material examined | protolog: AUSTRALIA: WESTERN AUSTRALIA—Shire of Dundaragan, Lesueur Nat. Pk., 6.vi.2010 E. M. & P. J. N. Davison EMD 14-2010 (holotype, PERTH 08351325). Localities withheld for conservation reasons - 5.vi.2010 E. M. & P. J. N. Davison EMD 9-2010 (paratype, PERTH 08351317), EMD 10-2010 (paratype, PERTH 08351295), 6.vi.2010 E. M. & P. J. N. Davison EMD 13-2010 (paratype, PERTH), 15.ix.2008 K. Syme KS 2110 (paratype, PERTH). | ||||||||||||
| discussion | |||||||||||||
| citations | —E. M. Davison and R. E. Tulloss | ||||||||||||
| editors | RET | ||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.

