


| name | Amanita griseibrunnea |
| name status | nomen acceptum |
| author | O. K. Mill. |
| english name | "Perth Lepidella" |
| intro |
The following is largely based on the original description of Miller (1992). |
| cap |
The cap of Amanita griseibrunnea is 30 - 65 (-83) mm wide, broadly convex to plane in age, orange-gray, darkening to brownish-gray, with a nonstriate and appendiculate margin. The volval remnants are present as white, mealy, slightly sticky volva, usually embedded with sand grains, and wearing away with age. The flesh is white, firm, and unchanging when bruised. |
| gills |
The gills are close, medium broad, narrowly adnate, white; short gills are present in at least one tier. |
| stem |
The stem is 60 - 130 × 11 - 23 mm, flaring somewhat at the top, otherwise cylindric, white, with a mealy-fibrillose powdery covering and irregular cottony patches of volva on the upper bulb. The bulb is narrow and short, sometimes rooting and usually covered with sand. The ring is in the upper part of the stem, white, cottony, mostly pressed to the surface of the stem forming an irregular zone, disappearing in older specimens. Before the ring separates from the cap margin, it forms a white, submembranous layer with soft, white scales arranged like spokes of a wheel on the bottom side. The flesh is white, firm, and unchanging when bruised. |
| odor/taste |
In young material, there is a faint odor of chlorine that becomes disagreeable in age. |
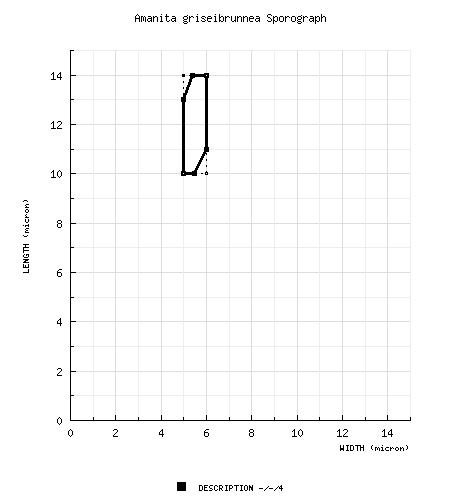
| spores | The spores measure (9.0-) 10 - 14 × 5 - 6 (-7) µm and are elongate to cylindric and amyloid. Miller observes that the amyloid reaction was unusually purple. Clamps are absent at bases of basidia. |
| discussion |
Amanita griseibrunnea was described from the state of West Australia. This species occurs in sandy soil under Eucalyptus marginata and a European imported pine (Pinus pinaster). While the pine was nearby, the present species was always found close to or in the root zone of Eucalyptus. This species was placed in section Lepidella by its author; and, given the description, we can further say it belongs in subsection Solitariae and further within the distinctive Australian stirps Straminea. This is due to the combination of a white, floccose and disappearing volva, an immarginate bulb at the stem base, and the lack of clamps in the fruiting bodies' tissue.—R. E. Tulloss and L. Possiel |
| brief editors | RET |
| name | Amanita griseibrunnea | ||||||||
| author | O. K. Mill. 1992a ["1991"]. Canad. J. Bot. 69: 2693, figs. 1-4. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Perth Lepidella" | ||||||||
| MycoBank nos. | 358164 [as griseobrunnea] | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | PERTH 02224518; isotype, VPI -> NY | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived entirely from the protolog of the present taxon. from protolog: Basidiomes are medium-sized. | ||||||||
| pileus | from protolog: 30 - 65 (- 83) mm wide, at first orange-gray (5B2), darkening to brownish gray (5B2 or 5C2), broadly convex to plane in age; context white, firm, unchanging when bruised; margin nonstriate, sometimes[?] appendiculate; universal veil initially as covering of white farinose "slightly sticky" material, usually embedded with sand grains, detersile. | ||||||||
| lamellae | from protolog: narrowly adnate, close, white, medium broad; lamellulae in one tier. | ||||||||
| stipe | from protolog: 60 - 130 mm × 11 - 23 mm, white, subcylindric, slightly flaring at apex; bulb “small” or as short rooting base, sand-covered; context as in pileus; partial veil in young specimens as intact submembranous veil with spoke-like soft white scales symmetrically arranged in radial pattern over underside, later as irregular superior annular zone of white appressed cottony material, prominent at first but hard to see or disappearing in older specimens; universal veil as white farinose to fibrillose or cottony irregular patches on upper part of bulb. | ||||||||
| odor/taste | from protolog: Odor when young faintly of chlorine, becoming stronger and disagreeable in age. Taste not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis |
Note: The original description does not appear to us to describe a typical pileipellis in Amanita. While there are a few cases of intercalary inflated segments in hyphae of the pileipellis in a few species, there is no other known report of pileocystidia in the genus. Since the universal veil is described as a covering of the pileus, it is most probable that the universal veil long remains attached to the pileipellis by abundant interconnecting hyphae. A re-examination of the anatomy of this species is required. Under these circumstances, even an apparently bare portion of the pileus surface will probably include inflated cells of the universal veil. We do not see a purpose in attempting to convert Miller's description to the format preferred by the editor.—ed. from protolog: up to 170 - 180 µm thick, as "loose to tangled trichodermium of cystidial end cells 62 - 100 × 10 - 22.5 µm, narrowly clavate, fusiform to cylindric, thin-walled, with filamentous to somewhat swollen hyphae 4 - 14 (-20) µm diameter, hyaline." | ||||||||
| pileus context | from protolog: filamentous hyphae 4 - 10 µm wide, mostly thin-walled, interwoven; acrophysalides not described; clamps absent. | ||||||||
| lamella trama | from protolog: bilateral, divergent; filamentous hyphae 4 - 10 µm wide, thin-walled, hyaline to yellowish in KOH; inflated cells not described; clamps absent. | ||||||||
| subhymenium | from protolog: narrow, cellular; inflated cells hyaline, isodiametric, 4 - 9 µm wide; clamps absent. | ||||||||
| basidia | from protolog: 36 - 48 × 9 - 11 µm, 4-sterigmate; clamps absent. | ||||||||
| universal veil | from protolog: On pileus[?]: filamentous hyphae 5 - 10 µm wide, sparse, thin-walled, branched; inflated cells numerous, (15 -) 22 - 55 µm wide, subglobose to pyriform to clavate, thin-walled, hyaline; clamps absent. | ||||||||
| stipe context | not described in protolog. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | from protolog: inflated cells hyaline, thin-walled, ovoid to pyriform, 9 - 15 × 12 - 24 µm. [Note: Originally misdescribed by Miller as "cheilocystidia." Perhaps Miller's additional phrase "scattered among the basidia" is a result of his seeing only scattered remnants of the largely deciduous, but originally continuous, lamella edge tissue.—ed.] | ||||||||
| basidiospores | from protolog: [-/-/4] (9.0-) 10.0 - 14.0 × 5.0 - 6.0 (-7.0) μm, (Q = 1.83 - 2.60; Q' = 2.17), thin-walled. amyloid (purple), elongate to cylindric; apiculus "small"; contents not recorded; pure white in deposit. | ||||||||
| ecology | from protolog: Rarely in groups of more than two. "Terrestrial, in loose sandy soil under or near Eucalyptus marginata with [alien] Pinus pinaster nearby." | ||||||||
| material examined |
from protolog:AUSTRALIA: WESTERN AUSTRALIA—City of Melville - Murdoch Univ. campus, buffer strip [32° 03' S, 115° 50' E], 7.v.1989 E. & P. Davison & O. K. & H. H. Miller [O. K. Miller] 23623 [E4273] (paratype, VPI), 7.vi.1989 O. K. Miller 23629 [E506] (holotype, PERTH; isotype, VPI). 9.vi.1991 E. Davison, L. & M. Bailey & O. K. & H. H. Miller [O. K. Miller] 24807 (paratype, VPI).
City of Perth - Kings Park, Additional material in VPI: AUSTRALIA: WESTERN AUSTRALIA—City of Melville - Murdoch Univ. campus[32°4'27" S/ 115°50'14" E], 18.vi.1989 Dr. Elaine Davison 8919 (VPI). | ||||||||
| discussion | from protolog: "Several of the collections were made in an area with [alien] P. pinaster nearby, but all collections were found close to the root zone of Eucalyptus. Sand often covers the pileus." | ||||||||
| citations |
The editors are grateful to Thomas Wieboldt, Curator of VPI, for his assistance in developing an accurate representation of collection data for extant original material of this species deposited in VPI. The editors express their thanks to Dr. Elaine Davison for her assistance with Western Australia geographical and other data relating to Miller's original materials of this taxon. —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita griseibrunnea |
| bottom links |
[ Section Lepidella page. ]
[ Amanita Studies home. ]
[ Keys & Checklists ] |
| name | Amanita griseibrunnea |
| bottom links |
[ Section Lepidella page. ]
[ Amanita Studies home. ]
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.