


| name | Amanita dryophila |
| name status | nomen acceptum |
| author | Consiglio & Contu |
| english name | "Dryad's Ringless Amanita" |
| synonyms |
=Amanita pianei Migl. & Lavorato nom. prov. |
| images |
 1. Amanita dryophila, Italy.  2. Amanita dryophila, Italy. |
| intro |
The following description is based on the original description by Consiglio and Contu (1999). |
| cap |
The cap of A. dryophila is (50-) 85 - 150 (-200) mm wide, more or less saturated hazel-brown, sometimes with orange shades, never with gray tints, rather fleshy, bell shaped at first, then convex to plano-convex, hardly ever completely expanded, viscid when moist, with a striate margin [according to the illustration, barely striate at first]. The volva is absent or present as detersile, white to slightly ochraceous, small crumb-like remnants. The flesh is firm, white, and unchanging. |
| gills |
The gills are free, rather broad, rather thick, narrowed at the ends, white, with a white to pale hazel-orange edge. |
| stem |
The stem is (60-) 100 - 150 (-200) × 15 - 30 (-40) mm, stuffed, rather strong, cylindric or narrowing upward, dry, white, flecked with pale hazel or hazel-orange fibrils in zigzag bands, with the lower part of the stem white or whitish, often shaded ochre. A ring is absent. The volva is saccate but brittle, easily broken, often difficult to collect without breaking, sometimes remaining as a belt of tissue on the lower half of the stem. The flesh is firm, white, and unchanging. |
| odor/taste |
Odor and taste are indistinct. |
| spores |
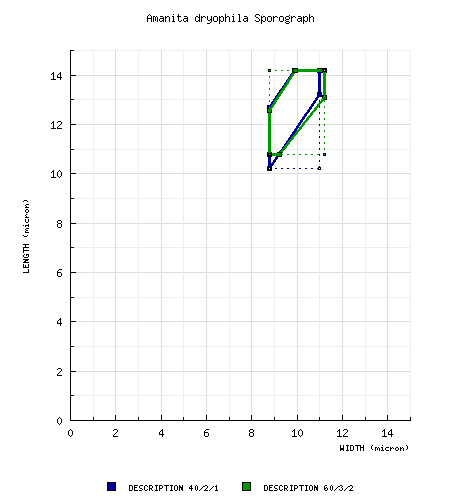
According to the original description, the spores measure 10.6 - 11.9 × 8.9 - 10.2 µm and are dominantly broadly ellipsoid, rarely ellipsoid or subglobose and inamyloid. Clamps are absent at bases of basidia. Spores from a paratype measured by RET yielded the following dimensions: (10.0-) 10.2 - 14.2 (-18.2) × (8.5-) 8.8 - 11.0 (-12.8) μm. |
| discussion |
This species was originally described from Italy but is also known from southern France. This species occurs in small groups in open sunny forests of broad-leaved trees, mainly oak (Quercus illex, Q. suber, Q. robur, etc.) on basic soil. In the original description, the discussion contains numerous errors. If section Vaginatae were to be divided into stirpes based on current knowledge, then the present species would indeed be placed with the group of species having broadly ellipsoid to ellipsoid spores, an obviously branching, largely uninflated subhymenium, and a fragile and not graying volva. In contradiction to the original description, the present species would be placed nearest Amanita malleata (Piane ex Bon) Contu and A. biovigera Singer. —R. E. Tulloss and L. Possiel |
| brief editors | RET |
| name | Amanita dryophila | ||||||||||||||||
| author | Consiglio & Contu. 1999. Persoonia 17: 287, 337 (pl. 15-16). [Note: pl. 16 depicts a collection with cap colors excluded by the protolog’s text.] | ||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||
| english name | "Dryad's Ringless Amanita" | ||||||||||||||||
| synonyms |
[=Amanita subavellanea Contu & Consiglio nom. prov. per Contu. 2000a. Boll. Gruppo Micol. G. Bresadola, N. Ser. 43(2): 239.]
=Amanita pianei Migl. & Lavorato nom. prov. 1987. Micol. Veg. Medit. 2(1): 43, unnumbered figs. p. 38.
?=Amanita inaurata var. alutacea E.-J. Gilbert. 1918. Gen. Amanita Pers.: 156. [The name is also ascribed to Bresadola or Quélet & Bataille, but neither proposed any name for the collection of Bresadola also cited by Gilbert. The name originates with Gilbert. Fraiture (1993. Op. Bot. Belg. 5: 87) suggests that this may be a synonym of Amanita mairei; however, the present taxon was originally described as having a copper colored pileus.] ≡Amanita vaginata var. alutacea (E.-J. Gilbert) Veselý. 1933. Ann. Mycol. 31(4): 289. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||
| MycoBank nos. | 460814 | ||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||
| holotypes | MCVE | ||||||||||||||||
| revisions | Migliozzi & Donato. 2022. Amanita section Vaginatae in Lazio. Second contribution. Taxonomic notes on Amanita dryophila. Mycol. Obs. 5: 57-71. [With correction of accession number of holotype, nrITS sequence from holotype, and plentiful illustrations.—ed.] | ||||||||||||||||
| selected illustrations | [?] Consiglio. 2000. Boll. Gruppo Micol. G. Bresadola 43(2): 226. [This image is a reproduction of the apparently nonconformant pl. 16 of the protolog. The pilei in the photograph have no hint of orange in the caps whatever, they appear to be sordid yellowish tan—at least as the photograph is reproduced. Photographic reproduction seems to RET to be of quite good quality in the journal's number in question. Therefore, it seems unlikely that the cluster of basidiomes in the photograph belong to the present species.] | ||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon (here presented in quotation marks) is based upon original research by R. E. Tulloss. [RET's note: Notes on a duplicate of a paratype collection were made 3-6 April 1993, a few months prior to RET's first efforts at defining biometric variables for use in describing the lamella trama and subhymenium; hence, such valuable information is missing from the account of the collection Migliozzi F0510A that is repeatedly cited below.] | ||||||||||||||||
| pileus |
"(50-) 85 - 150 (-200) mm broad, rather fleshy, convex to plano-convex, hardly ever completely expanded [in material seen by the authors—RET], with a striate margin, typically marked by some dimples and hollows, suggesting a semi-hypogeous development of the fruiting bodies, lubricous when wet, smooth, more or less saturated hazel-brown, sometimes with orange shades, never with grey tinges, often with detersile, white to slightly ochraceous small crumb-like volval remnants or glabrous." [RET's note: The authors are raising again an hypothesis already thoroughly debunked in the case of A. malleata—namely, that dimples on the pileus of an Amanita are an argument for hypogeous development because the dimples could be caused by pebbles. Such a thing is possible, but dimples can be seen in the caps of many common Amanita species that are well-known not to be hypogeous (e.g., Amanita muscaria and Amanita rubescens) and are much more likely to have another cause such as the collapse of tissue under the pileipellis when an insect larva begins to eat the tissue of the fungus after hatching from its egg.] | ||||||||||||||||
| lamellae | "rather broad and thick, ventricose, free, white with edge concolorous to pale hazel-orange, finely eroded." | ||||||||||||||||
| stipe |
"(60-) 100 - 150 (-200) × 15 - 30 (-40) mm, stuffed, rather strong, slender, cylindrical, progressively enlarged toward base, but never bulbous, dry, white, flecked with pale hazel or hazel-orange zig-zag bands. Partial veil strongly reduced, not forming an annulus. Universal veil brittle, friable, almost never integral but tending to remain in the ground, sometimes forming a sort of volval belt on lower half of stem, white or whitish, often shaded with ochre, not turning grey. Flesh firm, solid but strongly perishable, white unchanging." [RET's note: The last comment seems very odd. If there is no annulus, how can there be a partial veil when the two words are synonyms?] from revision of original material by Tulloss: universal veil as fragile saccate volva enclosing stipe base, whitish at first, with upper limb notably grayish or brownish in exsiccata, difficult to extract [in its entirety] from substrate. | ||||||||||||||||
| odor/taste | "Smell and taste indistinct." | ||||||||||||||||
| macrochemical tests |
none recorded. | ||||||||||||||||
| pileipellis |
"with an upper gelatinized region with cylindrical, very frail and irregularly arranged hyphae and with a lower region with cylindrical 3 - 8 μm wide, somewhat interwoven hyphae; ...." from revision of original material by Tulloss: extensively gelatinized at surface, orangish-yellow, 65 - 95 μm thick; filamentous undifferentiated hyphae subradially oriented, 3.5 - 5.0 μm wide, gelatinized; vascular hyphae 4,0 - 8.5 μm wide, common, branching, sinuous, without dominant orientation. | ||||||||||||||||
| pileus context |
"with some vascular hyphae." from revision of original material by Tulloss: orange-yellow for some distance below pileipellis; filamentous undifferentiated hyphae 1.0 - 10.8 μm wide, branching, loosely interwoven, in fascicles or not, plentiful, dominating near pileipellis; acrophysalides ovoid to broadly clavate to clavate to narrowly clavate to subfusiform (e.g., 66-78 × 26 - 33 μm), apparently subradially arranged, dominating except near pileipellis; vascular hyphae 3.5 - 8.0 μm wide, sinuous, common near interface with pileipellis, infrequent elsewhere, sometimes coiling or in loose tangles. | ||||||||||||||||
| lamella trama |
"bilateral." from revision of original material by Tulloss: bilateral; filamentous undifferentiated hyphae 2.8 - 8.5 μm wide, sometimes slightly constricted at septa, tangled and interwoven, frequently branching; terminal inflated cells not observed; vascular hyphae 2.8 - 6.0 μm wide, branching, locally tangled, in general not common; clamps not observed. | ||||||||||||||||
| subhymenium |
"composed of branching hyphae with elongate cells." [RET's note: Unfortunately there is no illustration to help the reader interpret the brief description.] from revision of original material by Tulloss: a frequently branching structure of largely uninflated hyphal segments (many rather short), with some intercalary clavate cells (e.g., 21 × 12.5 μm) in branching structure, with elements 1-3 cells below bases of basidia perpendicular to central stratum, with some hyphal segments parallel to central stratum, with basidia commonly arising from short uninflated hyphal segments. | ||||||||||||||||
| basidia |
"40 - 60 × 12 - 15 μm, 4-spored, rarely 2-spored, clavate, without clamps; ...." from revision of original material by Tulloss: 56 - 72 × 10.8 - 16.2 μm, about evenly divided between 4- and 2-sterigmate; clamps not observed. | ||||||||||||||||
| universal veil |
"with a subcellular texture consisting of cylindrical, 2 - 6 μm wide hyphae, and several inflated, 20 - 60 μm wide, hyaline to slightly ochraceous cells. Vascular hyphae numerous. Clamps absent." from revision of original material by Tulloss: On the stipe base, exterior layer (from upper, discolored region of limb): at least partially gelatinized, yellow-orange, exclusively comprising disoriented, interwoven filamentous undifferentiated hyphae, with some having slightly refractive yellowish walls, tangled or coiled or coparallel in fascicles; vascular hyphae not observed. On the stipe base, interior (in part of limb appressed to stipe): dominated by interwoven open matrix of hyphae; filamentous undifferentiated hyphae 3.0 - 11.5 μm wide, with somewhat thickened walls, often in fascicles, with some intercalary segments up to 18.0 μm wide; inflated cells clavate (up to 54 × 30 μm) to broadly ellipsoid (up to 54 × 43 μm) to subfusiform (e.g., 40 × 20 μm), with walls 0.5 - 0.8 μm thick, sometimes clustered, else scattered; vascular hyphae not observed. On the stipe base, interior (in upper, discolored portion of limb): inflated cells shaped as immediately above with addition of ovoid and subglobose cells, faintly sordid, with walls (especially of larger cells) up to 0.8 μm thick, up to 89 × 45 μm. On the stipe base, inner surface: gelatinization of surface or entire layer?, shallow layer of sublongitudinally oriented, gelatinization of surface or entire layer?, tightly interwoven filamentous undifferentiated hyphae. | ||||||||||||||||
| stipe context |
[Note: No description was provided in the
protolog. To formally complete the
identification of a specimen as an Amanita, it
is necessary to provide a description of the
longitudinally acrophysalidic tissue of the
stipe.—ed.] from revision of original material by RET: longitudinally acrophysalidic; filamentous undifferentiated hyphae 1.5 - 8.5 μm wide, branching, plentiful; acrophysalides up to 242 × 40 μm, plentiful; vascular hyphae 9.5 - 14.2 μm wide, rare, with slightly thickened walls. | ||||||||||||||||
| partial veil | absent. | ||||||||||||||||
| lamella edge tissue |
"25 - 60 × 12 - 30 μm, pyriform,
clavate, or sphaeropedunculate,
thin-walled." [Note: The cells on the lamella margin are of taxonomic value in some species. RET's current view is that it makes their description more valuable to describe the entire specialized tissue of which they are a part in addition to describing individual cell shapes and sizes.—ed.] | ||||||||||||||||
| basidiospores |
"10.6 - 11.9 × 8.9 - 10.2 μm
(average 11.3 × 9.6 μm), Q = 1.12 - 1.24 (average
1.18), hyaline, non-amyloid, broadly ellipsoid, rarely
ellipsoid or subglobose, sometimes lacrymoid, with a
large guttule inside, with a marked
apiculus." "Spore print
whitish." [Note: The sample size is not provided. It can be assumed to be rather small because of the very limited ranges given for length, width, and Q. There are eight collections cited in the protolog as material examined. Since the species occurs in a manner described as gregarious, these collections should contain more than one specimen each. It appears possible that spores were not measured from some of the specimens cited. The fact that herbaria of deposit are not cited for 6 of the eight collections makes an attempt at replicating the authors' results difficult.—ed.] from original material revised by Tulloss: [40/2/1] (10.0-) 10.2 - 14.2 (-18.2) × (8.5-) 8.8 - 11.0 (-12.8) μm, (L = 12.5 - 13.2 μm; L' = 12.8 μm; W = 9.7 - 10.0 μm; W' = 9.9 μm; Q = (1.16-) 1.20 - 1.44 (-1.48); Q = 1.29 - 1.32; Q' = 1.30), hyaline, colorless, smooth, thin-walled, ?amyloidity?, broadly ellipsoid to ellipsoid, adaxially flattened; apiculus sublateral, cylindric; contents monoguttulate; color in deposit not recorded. composite of all spore data from material revised by Tulloss: [60/3/2] (10.0-) 10.8 - 14.2 (-18.0) × (8.5-) 8.8 - 11.2 (-13.0) μm, (L = 12.2 - 13.2 μm; L' = 12.6 μm; W = 9.7 - 10.0 μm; W' = 9.9 μm; Q = (1.11-) 1.17 - 1.43 (-1.47); Q = 1.23 - 1.32; Q' = 1.28), hyaline, colorless, smooth, thin-walled, inamyloid, broadly ellipsoid to ellipsoid, infrequently subglobose, usually adaxially flattened; apiculus sublateral, truncate-conic to cylindric; contents monoguttulate; color in deposit not recorded. | ||||||||||||||||
| ecology |
Italy: "Terrestrial. gregarious
in open, sunny forests of broad-leaved trees,
mainly Quercus spp., on calcareous soil, often
bursting out of the soil. Autumn.
Not rare in Italy." [Note: Despite the description of the taxon as occurring in Autumn, five of the eight cited collections were made in the months of May, June, and July.—ed.] | ||||||||||||||||
| material examined |
"ITALY: BRESCIA—Bovegno, 1.x.1984 G. Guarinoni, A. Picena & G. Bontempi s.n. (no herb. cited, as "Amanita beckeri Huijsman"). EMILIA ROMANA—Prov. Reggio Emilia - Parco di Roncolo, 21.vi.1997 G Simonini s.n. (no herb. cited); Pulpiano, Viano, 22.vi.1997 G. Simonini s.n. (no herb. cited). Prov. Bologna - Monterenzio, Ronchi, 30.vi.1994 Consiglio 97020 (holotype, MCVE 736), 5.vii.1997 G. Consiglio 94025 (no herb. cited). Prov. Modena - Montese, selva di Castellauccio, s.d. G. Consiglio & E. Franceschini 94033 (no herb. cited).
LAZIO—Prov. Roma - Villa Pamphili, 10.v.1986 V. Migliozzi (CAG 2/1.30, fragment).
SARDEGNA—Prov. Nuoro - Laconi, loc. S. Sofia, 8.xi.1987 D. Farci & M. Rubia s.n. [M. Contu 87081101] (no herb. cited)." revised by Tulloss: FRANCE: GIRONDE—Gradignan-Mandavit, | ||||||||||||||||
| discussion |
RET is concerned that there may be typographical errors in the designation of the holotype and/or other material cited as reviewed in the protolog. It seems unusual that the collection number for material collected in 1994 should begin "97..." and the collection number for material collected in 1997 should begin "94..." So that the reader may compare the data, a description of A. pianei Migl. & Lavorato nom. prov. is included in the above data fields related to microscopic anatomy. It is based on a collection cited by Migliozzi and Lavorato (1987). A duplicate in CAG is cited as a paratype of A. dryophila, above. | ||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||
| editors | RET | ||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita dryophila |
| bottom links | [ Keys & Checklists ] |
| name | Amanita dryophila |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.