


| name | Amanita borneensis |
| name status | nomen acceptum |
| author | Boedijn |
| english name | "Borneo Lepidella" |
| images |
 1. Amanita borneensis, after a drawing of Boedijn, Borneo, Indonesia. |
| intro | The description of this species is based on that of Bas (1969). |
| cap | The cap of Amanita borneensis is about 60 - 130 mm wide, convex to plano-convex, rather fleshy, unequally pinkish brown, with a nonsulcate, strongly projecting, fuzzy margin. The cap is abundant with flat, rather small, brown, crust-like patches of volva. |
| gills | The gills are crowded, free to narrowly adnate, rather narrow, and pink. The short gills are truncate to obliquely truncate to attenuate. |
| stem | The stem is 150 - 200 × 15 - 25 mm, attenuate upward, solid, concolorous with cap or slightly paler, exannulate, with brown, rahter coarse wart-like remnants of volva near the base and on the upper part of the bulb. |
| spores | The spores measure 8 - 10 × 7.5 - 10 µm and are amyloid and globose to subglobose. Clamps are absent at bases of basidia. |
| discussion |
Amanita borneensis was described from Indonesia without mention of possible symbionts. Bas placed this species in his stirps Eriophora (see A. eriophora (Berk.) E.-J. Gilbert).—R. E. Tulloss |
| brief editors | RET |
| name | Amanita borneensis | ||||||||
| author | Boedijn. 1951. Sydowia 5: 324, fig. 4(1). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Borneo Lepidella" | ||||||||
| MycoBank nos. | 292444 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| lectotypes | BO (in liquid) | ||||||||
| lectotypifications | Bas. 1969. Persoonia 5: 478. | ||||||||
| type studies | Bas. 1969. Persoonia 5: 477, figs. 233-237. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The data below is derived from the study of the lectotype by Bas (1969). Bas (1969): Basidiome medium to large, rather slender. Yellow-brown granular vacuolar pigment present in all parts of basidiome. | ||||||||
| pileus | Bas (1969): 60 - 130 mm wide, "unequally pinkish brown" (between Ridgway's Russet Vinaceous and Sorghum Brown; but see discussion), convex to plano-convex, shiny where exposed; context whitish, rather fleshy; margin non-sulcate, strongly projecting, "fuzzy"; universal veil as abundant patches, flat, rather small, brown, crust-like. | ||||||||
| lamellae | Bas (1969): free to narrowly adnate, crowded, pink (more or less Ridgway's 'buff pink' or slightly paler), 4 -6 mm wide, with entire edge; lamellulae truncate to obliquely truncate (the shortest) to attenuate (the longer). | ||||||||
| stipe | Bas (1969): 150 - 200 × 15 - 25 mm, concolorous with pileus, attenuate upward, flocculose at apex, fibrillose to fibrillose-squamulose; bulb more or less globose, 40 - 50 mm wide; context solid, whitish; exannulate; universal veil in rather coarse wart-like fragments near stipe base and on upper part of bulb, brown, arranged more or less in rings. | ||||||||
| odor/taste | not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | Bas (1969): gelatinized near surface except near margin; filamentous hyphae 2 - 15 μm wide, near margin radially oriented, otherwise interwoven, with brown vacuolar pigment; vascular hyphae scattered, brown; clamps absent. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | Bas (1969): bilateral; almost certainly with inflated terminal cells up to 110 × 30 μm and terminal rows of shorter cells; clamps absent. | ||||||||
| subhymenium | Bas (1969): rather broad, difficult to delimit, subcellular to cellular; clamps absent. | ||||||||
| basidia | Bas (1969): 45 - 60 × 11 - 15 μm, 4-sterigmate; clamps absent; "scattered basidium-like cells with one rather thick, cylindrical appendix occurring among normal basidia"; clamps absent. | ||||||||
| universal veil | Bas (1969): On pileus: elements irregularly disposed; filamentous hyphae 2 - 15 μm wide, rather abundant; inflated cells predominantly with brown granular contents, sometimes with walls thickened (especially near pileus margin), globose (up to 60 μm wide) and ellipsoid to ventricose-fusiform (up to 90 × 50 μm), predominantly terminal singly [sometimes subtended by ventricose to subcylindric cell (up to 200 μm long and 15 - 24 μm wide)], more rarely in short chains; clamps absent. In "fuzzy" fringe on pileus: filamentous hyphae dominant, loosely interwoven, subradially oriented, 10 - 20 μm wide; inflated cells cylindric to slenderly fusiform to fusiform, more rarely ellipsoid to globose, terminal singly or in short chains. On stipe base: elements irregularly disposed; filamentous hyphae 6 - 10 (-20) μm, abundant, often with dark brown contents, often with thickened walls; inflated cells scattered, mostly elongate-ellipsoid or fusiform, but also clavate or pyriform or ellipsoid, up to 200 × 65 μm; clamps absent. [Note: Bas segregated the "fuzzy" fringe of the pileus from the tissues he described as the universal veil; RET has chosen to include the information in the "universal veil" data field due to the similarity of the structure of the "fuzzy" fringe with the structures reported in warts on the pileus and the stipe base.—ed.] | ||||||||
| stipe context | Bas (1969): longitudinally acrophysalidic; filamentous hyphae 5 - 10 μm wide; acrophysalides 120 - 360 × 15 - 40 μm; clamps absent. | ||||||||
| partial veil | absent. | ||||||||
| lamella edge tissue | Bas (1969): as "rather thick rim"; filamentous hyphae 4 - 7 μm wide; inflated cells in irregularly disposed chains (10 - 35 μm wide, globose) or terminal singly (mostly pyriform, up to 50 × 35 μm). | ||||||||
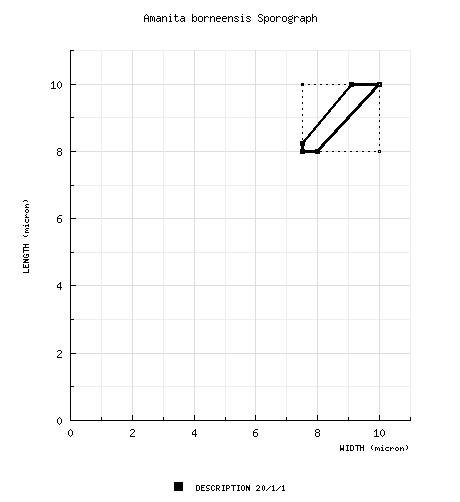
| basidiospores | Bas (1969): [20/1/1] 8.0 - 10.0 × 7.5 - 10.0 μm, (Q = 1.0 - 1.10; Q = 1.05), thin-walled, amyloid, globose to subglobose; apiculus proportionately small; contents granular, brownish-yellowish; color in deposit not recorded. | ||||||||
| ecology | Bas (1969): At ca. 700 m elev. Terrestrial. | ||||||||
| material examined |
Bas (1969): INDONESIA: BORNEO—Schwaner Gebergte, Bukit Bidang Menabai, ca. Bukit Raja, | ||||||||
| discussion |
Bas (1969): "Boedijn (1951) did not mention a number for the type collection. His drawing of A. borneensis in the Rijksherbarium at Leiden...[that is, the source for Bas' illustration used on this page—ed.] shows three fruit-bodies and bears the numbers 8936 and 8888, with no indication to which fruit-bodies the numbers belong. On the label of the collection received on loan from the herbarium at Bogor both the number 8936 and the indication 'Borneo, Dec. 1924' occur. This collection is selected as lectotype. The syntype BO 8888 has the following data on its label: 'Borneo 1924, Hans Winkler 1117'. It is kept in Bogor, preserved in liquid in a jar too large for shipping. "Amanita borneensis is a large species with pinkish brown cap and stem, a brown, friable volva leaving small patches on the cap and rather coarse warts at the base of the bulbous, exannulate stem, a fuzzy rim along the border of the cap, globose spores, and lacking clamps. "Because of these characters A. borneensis resembles strongly A. berkeleyi from India..., which unfortunately is rather imperfectly known. It is possible that in the future these two species will turn out to be conspecific, but for the moment A. borneensis is maintained as an independent species because of (i) the pinkish tinge in the brown colour of cap and stem, (ii) the pinkish gills, (iii) the more strictly globose to subglobose spores, and (iv) the larger inflated cells of the volva. "In connection with the mentioned difference in colour between A. berkeleyi and A. borneensis it is of interest that the pigment of the elements of the pileipellis and volva in the material of borneensis in alcohol preserved is very dark. That is why I should have expected much darker colours than those recorded in the original description. I have not seen the field-notes from which Boedijn must have taken his notes on the colours of the fruit-body. The preserved specimen is now entirely dark brown. "The volva of A. borneensis has been described as circumcised and in the drawings...a groove between stem and bulb is visible. However, such a groove is present only in one half of the bulb of the type specimen and certainly does not mark the upper limit of the remnants of the volva, which can still be found several centimeters above this groove. "A species closely related to A. borneensis has recently been collected by Prof. E. J. H. Corner on Mt. Kinabalu in North Borneo (A. ornatipes Bas ined.; RSNB 1756; L). It differs from A. borneensis in its grey cap and volva, white gills, and broadly ellipsoid to ellipsoid spores (9 - 11 × 6.5 - 8.5 μm, l/b 1.2 - 1.5)." | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita borneensis |
| name status | nomen acceptum |
| author | Boedijn |
| english name | "Borneo Lepidella" |
| images |
1. Amanita borneensis, after a drawing of Boedijn, Borneo, Indonesia. |
| drawing | Dr. C. Bas (1969) after the drawings of Boedijn, reproduced by courtesy of Persoonia, Leiden, the Netherlands |
| name | Amanita borneensis |
| bottom links | [ Keys & Checklists ] |
| name | Amanita borneensis |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.