


| name | Amanita annulatovaginata |
| name status | insufficiently known |
| author | Beeli |
| english name | "African Slender Caesar" |
| intro | The following description is based on the original description of Amanita annulatovaginata and (Gilbert 1940 & 1941). |
| cap | The cap of Amanita annulatovaginata is 80 - 90 mm wide, ovoid then plano-convex, slightly umbonate, umbrinous brown to fuligineous, and undecorated. The cap margin has long striations. The flesh is white and thin. |
| gills | The gills are free and white. The short gills have not been described. |
| stem | Its stem is 180 - 190 × 7 - 8 mm, cylindric, stuffed, undecorated, bistre, and brownish-black. The ring is superior, submembranous, white, and easily broken. The volva is saccate, membranous, ample, and bistre or umber. The flesh is white. |
| odor/taste | Neither taste nor odor were recorded for this mushroom. |
| spores |
Spores from the type and a
another collection having a water color suggesting that it also belongs to the type variety are illustrated in (Gilbert 1940). These 9 spores are 11.1 - 14.9 × 6.5 - 10.5 µm and ellipsoid to elongate and inamyloid. The following data also comes from Gilbert's drawings: Spores from the type of A. annulatovaginata var. atra are 9.9 - 11.1 × 6.6 - 7.8 µm [5 spores] and ellipsoid. Spores from var. citrina are 12.3 - 12.9 × 7.9 - 8.3 µm [2 spores] and ellipsoid to elongate. Spores identified only at the species rank, but not the same size as those of the type are 10.3 - 11.9 × 6.0 - 7.7 µm [4 spores] and are ellipsoid to elongate. |
| discussion |
The present species was originally described from the Democratic Republic of Congo in forest. Madame Goossens' watercolors show the stipe completely distinguishable from the volval sac and the annulus more or less persistent at maturity. Beeli distinguished four varieties within this species. In addition to the type variety, he proposed Gilbert (1940) proposed that var. atra and var. amethystina were essentially identical with the type variety. However, his argument is problematic. See the "discussion" data field of the technical tab of this page. The form of the fruiting bodies suggests placement of A. annulatovaginata and two of its varieties in stirps Hemibapha, but there is insufficient information to feel confident about this placement.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita annulatovaginata | ||||||||
| author | ("annulato-vaginata") Beeli. 1927. Bull. Soc. Roy. Bot. Belgique 59: 103, pl. I (fig. 5). | ||||||||
| name status | insufficiently known | ||||||||
| english name | "African Slender Caesar" | ||||||||
| synonyms |
≡Amanitopsis annulatovaginata (Beeli) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 76, tab. 8 (figs. 1-4). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 203645, 284112 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| lectotypes | BR | ||||||||
| lectotypifications | E.-J. Gilbert. 1940. op. cit.: 96 (in caption to tab. VIII (fig. 1)). | ||||||||
| selected illustrations |
Beeli. 1935. Fl. Iconogr. Champ. Congo 1: pl. II (figs. 2,4). E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl. (1): tab. 12 (fig. 1). [From same watercolor as in Beeli (1935).] | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present taxon and Gilbert (1940 & 1941). | ||||||||
| pileus | from protolog: 80 - 90 m wide, umbrinous brown to fuligineous, ovoid then plano-convex, umbonate, glabrous; context thin; margin long-striate (ca. 0.3-0.4R per figure); universal veil often absent, sometimes as single membranous calyptra. | ||||||||
| lamellae | from protolog: free, density not recorded, white; lamellulae not described. | ||||||||
| stipe | from protolog: 180 - 190 × 7 - 8 μm, bister, cylindric; context hollow; partial veil superior, membranous, very friable, white; universal veil saccate, membranous, ample, bister or umber. | ||||||||
| odor/taste | not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | not described. | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | not described. | ||||||||
| subhymenium | not described. | ||||||||
| basidia | not described. | ||||||||
| universal veil | not described. | ||||||||
| stipe context | not described. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | not described. | ||||||||
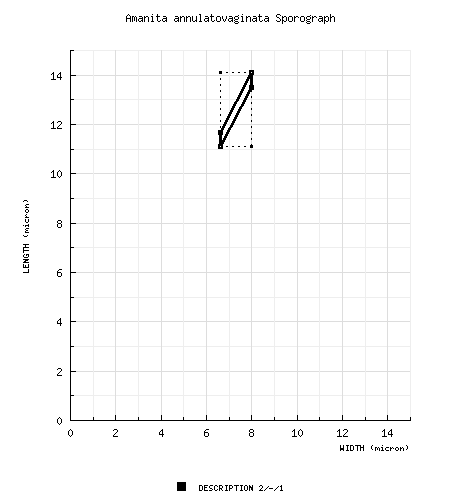
| basidiospores | Gilbert (1940), from lectotype: [2/-/1] 11.1 - 14.1 (-14.2) × 6.6 - 8.0 (-9.6) μm, (L = 12.6 μm; W = 7.3 μm; Q = 1.69 - 1.77; Q = 1.73. [Note: The spore measurements are taken from the two drawings of (Gilbert 1940: tab. VIII (fig. 1)) that are in apparent lateral view.—ed.] | ||||||||
| ecology | from protolog: Scattered. Terrestrial in dry forest. | ||||||||
| material examined |
from protolog: CONGO, DEMOCRATIC REPUBLIC OF: PROV. EQUATEUR—Territoire Mbandaka | ||||||||
| discussion |
Beeli reported an indigenous name for this species—"bempokna." Gilbert's actions—designating a lectotype for the present species in 1940, then proposing to designate the implicit hololotype (sole collection) of A. annulatovaginata var. atra as "lectotype" of the present species in 1941—are flawed. Whatever the condition of Goossens 251 (the material exists to this day and is supported by a watercolor), Gilbert ignored the second syntype (also supported by a watercolor); hence, there was ample original material from which to select a lectotype (and he had already done so). Gilbert emphasizes that the lectotype of the type variety is composed of old specimens in bad condition and that the sole collection (implicit holotype) of var. atra is, in contrast, a good, representative collection of the same taxonomic entity. This cannot be the case. Gilbert's proposal of relative age would mean that the spores of Goossens 251 would be the spores of an aging specimen; and, hence, should be shorter and, probably, of lower Q than those of Goossens 773 (if it were in the prime of sporulation); however, the exact opposite is the case. Moreover, Goossens' watercolor of var. atra [reproduced in both (Beeli 1935: pl. II (figs. 5a-c)) and (Gilbert 1941: tab. 11)] shows a flocculent upper stipe with no evidence of a true partial veil; and, from this evidence, it seems doubtful that var. atra is assignable to sect. Caesareae. Hence, we suggest treating all the varieties as separate taxa until we have a much better understanding of them, and var. atra has been moved provisionally to section Vaginatae on this site. | ||||||||
| citations |
—R. E. Tulloss and A. Fraiture [Note: The editors acknowledge the generous support of Dr. André Fraiture (BR) in assisting with the authorship of this page.—eds.] | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita annulatovaginata |
| bottom links |
[ Keys & Checklists ] [ sub-Sahara checklist ] [ Draft description of, & key to, sect. Caesareae ] |
| name | Amanita annulatovaginata |
| bottom links |
[ Keys & Checklists ] [ sub-Sahara checklist ] [ Draft description of, & key to, sect. Caesareae ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.