


| name | Amanita subcaligata |
| name status | nomen acceptum |
| author | (A. H. Sm. & P. M. Rea) A. H. Sm. ex Tulloss in Volk & Burds. |
| english name | "Salmon Lepidella" |
| synonyms |
=Amanita salmonea Thiers |
| images |
 1. Amanita subcaligata, Brazos Co., Texas, U.S.A.  2. Amanita subcaligata, Brazos Co., Texas, U.S.A. |
| cap | The cap of A. subcaligata is 50 - 110 mm wide, smooth and salmon-buff at first, soon areolate, conico-convex to convex to nearly plane, and occasionally with a slight umbo or with a slightly depressed center. The margin is nonstriate and slightly appendiculate. The volva covers the entire cap at first and is connected directly to the cap flesh—with no cap skin in between. As the cap expands the volval layer breaks up into subpyramidal regions about 1 - 5 mm wide and exposing tissue that may be more volva or cap flesh ranging in color from white to seashell pink to pale buff to buff (after some drying). Unexposed flesh is white when cut. As time passes, volval warts become pinkish brown to cinnamon-rufus to hazel with highest points darkest. The volval material is more fibrillose and appressed toward the cap margin. |
| gills | The gills are narrowly adnate to free, sometimes seceding and leaving striations on the stem's apical region, subdistant to crowded, cream in mass, in side view pale cream to off-white to white with a faintly yellowish and "waxy" tinge; they are 6 - 18 mm broad. When the gills fork (not often seen), this happens close to the stem. Short gills are truncate to subtruncate to attenuate, unevenly distributed, of diverse lengths, and plentiful. |
| stem | The stem is about 60 - 127 × 8 - 20 mm, white to off-white, becoming pale brown from handling, cylindric or narrowing downward, occasionally slightly expanded at the base, sometimes slightly dog-legged, and occasionally with a rounded point at the very bottom. The stem does not flare at the apex; it is glabrous to subfibrillose and very finely longitudinally striate above the annulus. The context is white and unchanging or faintly buff in the base and sometimes tan near stained surface regions. The stem bears an annulus that is superior, skirt-like, membranous, rather narrow, bearing a thickened edge, and cream at first, but then taking on a pale brown tint. The lower surface of the annulus is cottony-floccose. The thickest scales on the bottom of the annulus become brown on the tips as the volva does on the cap. The annulus eventually tears and collapses and may be lost. The volva is distributed from just below the annulus nearly to the stem base in roughly concentric scales or (especially lower on the stem and within the ground) as nearly complete rings (suggesting the volva at the top of the bulb in A. muscaria (L. : Fr.) Lam.). Some rings may be at a pronounced angle to the level of the ground. The volval remnants are pallid to yellowish brown to rusty orange (a very striking color that I have seen on one specimen of the present species in Colorado) to brown on the exterior, but white within. |
| spores | The spores measure (8.5-) 9.3 - 13.0 (-17.7) × (6.2-) 6.5 - 10.2 (-11.5) >µm and are amyloid and broadly ellipsoid to ellipsoid. The spore print is white. Clamps are plentiful and rather large at bases of basidia. |
| discussion |
The odor is indistinct at first, but may become foetid or "like dog urine." The taste is indistinct. Since some taxa of subsection Vittadiniae are POISONOUS, eating A. subcaligata is not recommended. Amanita subcaligata was described originally in Armillaria from "under shrubs" from the state of California, U.S.A. Under the synonym, A. salmonea, this species was described from fairy rings in lawns in Texas. The species is also known from high altitude prairie and grazing land in Colorado. In Texas, it sometimes is found intermixed with A. silvifuga Bas. Reports of A. subcaligata from Europe have proven to be false. Recent placement of the species in Floccularia was also incorrect. It is without doubt an Amanita.< Bas (1969) placed A. subcaligata (as "A. salmonea") in his stirps Vittadinii, in which he noted that the present species was most similar to A. ameghinoi (Speg.) Singer, known only from Argentina.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita subcaligata | ||||||||||||||||||||||||||||||||||||
| author | (A. H. Sm. & P. M. Rea) A. H. Sm. ex Tulloss in Volk & Burds. 1995. Synopsis Fung. (Oslo) 8: 112. | ||||||||||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||||||||||
| english name | "Salmon Lepidella" | ||||||||||||||||||||||||||||||||||||
| synonyms |
≡Armillaria subcaligata A. H. Sm. & P. M. Rea. 1944. Mycologia 36: 128.
≡Amanita subcaligata (A. H. Sm. & P. M. Rea) R. Henry nom. inval. 1961. Rev. Mycol. (Paris) 36: 223. [Lacking full and direct reference to basionym. ICBN §33.2.]
≡Floccularia subcaligata (A. H. Sm. & P. M. Rea) Bon. 1991. Doc. Mycol. 21(81): 55.
=Amanita salmonea Thiers. 1957. Mycologia 49: 720. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||||||||||||||||||
| MycoBank nos. | 434357 | ||||||||||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||||||||||
| holotypes | Armillaria subcaligata & Amanita salmonea—MICH | ||||||||||||||||||||||||||||||||||||
| revisions |
A. salmonea—Bas. 1969. Persoonia 5: 360, figs. 45-47. A. subcaligata—Tulloss, here | ||||||||||||||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly the protologs is based on original research by R. E. Tulloss. | ||||||||||||||||||||||||||||||||||||
| pileus | 50 - 110 mm wide, smooth and Salmon-Buff (7.5YR 7.8/6.0) at first, convex to conico-convex to broadly convex to nearly plane, sometimes eventually plano-concave, occasionally with broad umbo or slight central depression, dry, soon areolate and sometimes strongly so (breaking up into subpyramidal regions 1 - 5± mm wide) exposing white to Seashell Pink (7.5YR 8.0/3.0) to pale buff or buff (after some in situ drying) context, dry, matt to subshiny (after some in situ drying); context pure white in tissue not exposed on surface, soft, 9.5 - 13 mm thick at stipe, thinning evenly to margin and there about 2 mm thick, then continuing to thin evenly to edge of sterile rim, not changing color when bruised or cut; margin nonstriate, with sterile rim striate on inner surface and extending 3 - 4 mm beyond lamellae, at least sometimes with 1 mm wide sinus between ends of lamellae and sterile rim; universal veil as upper portion of large subpyramidal warts, becoming pinkish brown to Cinnamon-Rufus (2.5YR 5.0/8.0) to Hazel (2.5YR 4.4/5.5) with highest points darker, adnate, becoming more fibrillose and appressed toward margin. | ||||||||||||||||||||||||||||||||||||
| lamellae | narrowly adnate to free, seceding, leaving striations on stipe apex, crowded to subdistant, cream in mass, pale cream to off-white to white with faintly yellowish and “waxy” tinge, 6 - 18 mm broad, thin, with edges entire to eroded, not forking or occasionally forking very close to stipe; lamellulae truncate to subtruncate to attenuate, unevenly distributed, of diverse lengths, plentiful. | ||||||||||||||||||||||||||||||||||||
| stipe | 60 - 127 × 8 - 20 mm, cylindric or narrowing downward, occasionally slightly enlarged at base, sometimes slightly dog-legged, occasionally with rounded point at base, white to off-white, becoming pale brown from handling, subcylindric or narrowing upward, not flaring at apex, glabrous to subfibrillose and finely striatulate (10× lens) above partial veil, fibrillose subsilky and longitudinally striatulate below; context white, unchanging or faintly buff in very base, sometimes tan near stained regions on surface, solid, with larval tunnels concolorous; partial veil superior, submembranous, patent to pendent, rather narrow, with thickened edge, 1± mm thick at first, cream at first, then taking on very pale brown tint, with upper surface smooth to very faintly striate and lower surface cottony-floccose with thick scales having brownish tips similar to warts on pileus, soon collapsing or tearing and becoming inconspicuous; universal veil distributed from just below annulus nearly to base of stipe in subconcentric scales, on lower stipe and within substrate as broken rings somewhat as in A. muscaria or as nearly complete rings at pronounced angle to substrate surface, pallid (especially on upper stipe at first) to yellowish brown to rusty orange to brown on exterior, but white within (hence, with surface color possibly due to oxidation or gelatinization). | ||||||||||||||||||||||||||||||||||||
| odor/taste | Odor indistinct (protolog of A. salmonea) or, sometimes after some in situ drying, foetid or like dog urine. Taste indistinct (protolog of A. salmonea). | ||||||||||||||||||||||||||||||||||||
| macrochemical tests |
Spot test for laccase (syringaldazine) - completely negative throughout basidiome. Spot test for tyrosinase (paracresol) - positive in context of pileus disc and context of central upper stipe, over much of lower stipe surface and in context at very base, in spots in pileus context just above lamellae. Test voucher: Tulloss 8-19-97-A. | ||||||||||||||||||||||||||||||||||||
| pileipellis | lacking. | ||||||||||||||||||||||||||||||||||||
| pileus context | filamentous, undifferentiated hyphae 2.4 - 11.9 µm wide, branching, loosely interwoven; acrophysalides plentiful to dominant, thin-walled, elongate to narrowly clavate to clavate to ovoid, to 191 × 77 µm; vascular hyphae 4.2 - 14.3 µm wide, branching. | ||||||||||||||||||||||||||||||||||||
| lamella trama | bilateral; wcs = 55 - 65 µm; central stratum apparently totally lacking inflated cells; filamentous, undifferentiated hyphae approaching subhymenium in disorderly manner via cluster of short branch segments, with cells of subhymenium arising from such branch segments; filamentous, undifferentiated hyphae 2.2 - 7.0 µm wide; vascular hyphae 3.0 - 7.0 µm wide, branching, sinuous; clamps present. | ||||||||||||||||||||||||||||||||||||
| subhymenium | ws-near = 50 - 55 µm; ws-far = 60 - 65 µm; comprising clavate to broadly clavate to ellipsoid inflated cells (up to 29 × 19 µm) and partially inflated hyphal segments in 4 - 5 layers, locally cellular, with occasional single large ovoid cell reaching from branching hypha of trama to base of basidium, with major diameter of many elements perpendicular to central stratum, with basidia arising singly or in pairs from cells of all types. | ||||||||||||||||||||||||||||||||||||
| basidia | 35 - 63 × (8.5-) 9.8 - 16.0 µm, thin-walled, many swollen to greatest width in region in which they pass beyond apices of surrounding basidioles, mostly 4-, and occasionally 2-, sterigmate, with sterigmata to 5.0 × 2.5 µm; clamps plentiful and often rather large. | ||||||||||||||||||||||||||||||||||||
| universal veil | On pileus: continuation of elements of pileus context without hiatus, in little space taking on anticlinal orientation~—perpendicular to similar elements of pileus context; filamentous, undifferentiated hyphae dominant, 2.1 - 9.8 µm wide; inflated cells thin-walled, rather narrow, vertically oriented, terminal or in brief chains, to 129 × 34 µm or larger; vascular hyphae 4.8 - 6.0 µm wide, especially notable near and in region of divergence of universal veil from pileus context; tissue yellow to orange-yellow in 3% KOH in regions of greatest gelatinization of warts; clamps present. Along stipe (below annulus): Dark, concentric, incomplete, wavy lines seemingly representing collapsed, gelatinized universal veil remnants, orange to orange-yellow when mounted in 3% KOH, appearing to be extensively gelatinized material with remnants of hyphae and possibly of inflated cells aligned in smoothly flowing curves following the ungelatinized elements from longitudinal orientation to transverse and back again. | ||||||||||||||||||||||||||||||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 3.0 - 10.5 µm wide, branching, plentiful to dominating; acrophysalides clavate, thin-walled, plentiful to locally dominating, to 213 × 45 µm; vascular hyphae 2.5 - 4.9 µm wide, branching; clamps common and moderately prominent. | ||||||||||||||||||||||||||||||||||||
| partial veil | extensively gelatinized especially on surfaces; filamentous, undifferentiated hyphae interwoven, 1.8 - 9.1 µm wide, collapsing and gelatinizing, often in subradially oriented fascicles; inflated cells collapsing, scattered, thin-walled, clavate, up to 96 × 29 µm; vascular hyphae scarce or absent; clamps present. | ||||||||||||||||||||||||||||||||||||
| lamella edge tissue | not recorded. | ||||||||||||||||||||||||||||||||||||
| basidiospores |
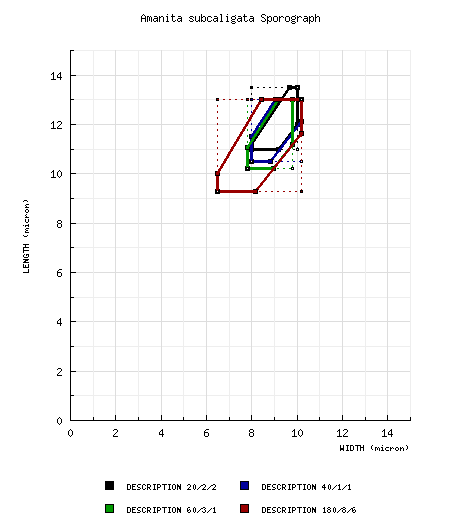
Bas (1969) as A. salmonea: [20/2/2] 11.0 - 13.5 × 8.0 - 10.0 μm, (Q = (1.10-) 1.20 - 1.40 (-1.55); Q = 1.30 - 1.35). RET type study of Armillaria subcaligata: [40/1/1] (8.8-) 10.5 - 13.0 (-14.5) × (7.8-) 8.0 - 10.2 (-11.5) μm, (L = 11.9 μm; W = 9.0 μm; Q = (1.13-) 1.19 - 1.44 (-1.48); Q = 1.32). RET type study of A. salmonea: [60/3/1] (9.2-) 10.2 - 13.0 (-14.0) × (7.0-) 7.8 - 9.8 (-11.2) μm, (L = 11.4 - 11.9 μm; L' = 11.6 μm; W = 8.7 - 9.1 μm; W' = 8.8 μm; Q = (1.09-) 1.14 - 1.42 (-1.47); Q = 1.31 -1.32; Q' = 1.32). composite of data from all material revised by RET: [180/8/6] (8.5-) 9.3 - 13.0 (-17.7) × (6.2-) 6.5 - 10.2 (-11.5) µm, (L = 9.9 - 11.9 (-12.4) µm; L’ = 11.4 µm; W = (6.7-) 8.0 - 9.5 µm; W’ = 8.6 µm; Q = (1.05-) 1.14 - 1.54 (-1.93); Q = 1.22 - 1.44 (-1.48); Q’ = 1.33), hyaline, colorless, smooth, thin-walled, amyloid, broadly ellipsoid to ellipsoid, occasionally subglobose, often adaxially flattened, occasionally as giant spores and then often of irregular shape; apiculus sublateral, truncate-conic; contents granular to guttulate; white in deposit. | ||||||||||||||||||||||||||||||||||||
| ecology | Solitary to gregarious. California: In dry lawn near shrubbery. Colorado: At 1715 m elev. In fine soil with high organic content, growing through pile of dead grass or straw at edge of drainage ditch with nearest ectomycorrhizal tree (Populus) at distance of 15 m and across ditch. Texas: At ca. 115 m elev. In fairy rings on lawns. | ||||||||||||||||||||||||||||||||||||
| material examined |
U.S.A.: CALIFORNIA—Santa Barbara Co. - Santa Barbara, 7.viii.1941 Heloise Coutelenc s.n. [P. M. & M. Rea 994] (holotype, MICH in two boxes; fragments in L).
COLORADO—Jefferson Co. - just W of Lakewood, Applewood Grove (uninc.), just off Willow Ln. [39°44’43” N/ 105°08’51” W], 19.viii.1997 Rosa-Lee and Robert Brace s.n. [Tulloss 8-19-97-A] (DBG 18895; RET 266-6).
TEXAS—Brazos Co. - College Station, 14.ix.1951 Harry D. Thiers 1393 (holotype of A. salmonea, MICH), 23.x.1957 H. D. Thiers 4701 (L n.v.); College Station, YMCA Blvd. [30°36.927’ N/ 96°20.485’ W, ca. 115 m], 16.x.1982 David P. Lewis 3335 (in herb. David T. Jenkins n.v.; RET 033-7; SFSU); College Station, Texas A. & M. Univ. campus, 6.x.1957 H. D. Thiers 4630 (SFSU as "A. monticulosa," mixed collection dominated by A. silvifuga); College Station, Texas A. & M. Univ. campus, ca. Ross statue, 16.x.1982 D. P. Lewis 3336 (RET 033-8). [NOTE: many E. A. Smith collections in MICH and much Thiers material in SFSU.] | ||||||||||||||||||||||||||||||||||||
| discussion |
[Note: There’s considerably more material in MICH. For example, A. H. S. s.n. 6.vi.1977 ("Amanita subcaligata part to Bas") and many collections of A. salmonea. Also, E. A. Smith colls. in MICH marked "A. praelatifolia." Also (ENCB), get Guzmán 6831 from Hidalgo (SW of Huasca, 28.vii.1968).] For some time, it was recognized that Armillaria subcaligata is assignable to Amanita. A. H. Smith (1979) made this observation in his discussion of Armillaria stirps Caligata. Thiers (1982) kept this information easily accessible by including the protolog of Armillaria subcaligata in its entirety in his chapter on the Amanitaceae in his Agaricales in California project. Bas (1969) distinguishes Amanita salmonea and A. ameghinoi (Speg.) Singer (described from Argentina) from all other taxa in subsect. Vittadiniae because of the universal veil which he presumed to be turf-like in young basidiomes and continuous with the pileus context in both cases. As the pileus expands, the universal veil breaks up into subpyramidal warts with tips browning due to the gelatinization of the uppermost elements. Armillaria subcaligata exhibits this same unusual universal veil. Bas considered that A. salmonea and A. ameghinoi might be contaxic. In a collection of A. ameghinoi (BAFC 30.888) which is, in general, in much better condition than is the type of that species, I was unable to find any structure at the pileus surface that was not periclinally oriented; however, there was very little remaining of the universal veil. Bas’ report that the acrophysalides were scarce in the stipe context of A. salmonea was of sufficient concern that I reviewed carefully the holotype of that taxon. I found acrophysalides to be common in the portion of tissue I inspected; although it is certainly the case that filamentous, undifferentiated hyphae dominated the tissue. There is no significant difference in spore size or shape in the two holotypes: BASIDIOSPORES (from holotype of Amanita salmonea): [60/3/1] (9.2-) 10.2 - 13.0 (-14.0) × (7.0-) 7.8 - 9.8 (-11.2) µm, (L = 11.4 - 11.9 µm; L’ = 11.6 µm; W = 8.7 - 9.1 µm; W’ = 8.8 µm; Q = (1.09-) 1.14 - 1.42 (-1.47); Q = 1.31 - 1.32; Q’ = 1.32), hyaline, smooth, thin-walled, amyloid, ellipsoid to broadly ellipsoid, occasionally subglobose, often somewhat adaxially flattened, often expanded at one end; apiculus sublateral, truncate-conic; contents monoguttulate to guttulate to granular. BASIDIOSPORES (from holotype of Armillaria subcaligata): [40/1/1] (8.8-) 10.5 - 13.0 (-14.5) × (7.8-) 8.0 - 10.2 (-11.5) µm, (L = 11.9 µm; W = 9.0 µm; Q = (1.13-) 1.19 - 1.44 (-1.48); Q = 1.32), smooth, hyaline, thin-walled, amyloid, broadly ellipsoid to ellipsoid, occasionally subglobose, often adaxially flattened; apiculus sublateral, truncate-conic; contents guttulate to granular. Based on study of the two types, I propose that A. salmonea is a taxonomic synonym A. subcaligata. Like a number of the taxa in subsection Vittadinii, this species is extremely difficult to dry and keep dry. In correspondence with A. H. Smith recorded in Smith’s manuscript notes on Amanita, E. A. Smith wrote concerning a large collection of the present species made near Cisco, Texas that it was difficult to dry. In the same letter, E. A. Smith noted a tendency of the lamellae to "stain red" in drying and noted that in one collection "the gills were flushed greenish at first." For comparison with A. silvifuga, see the discussion following the description of that species. The material reported from Hidalgo, Mexico, by Guzmán (1975) was examined by me; and, as Guzmán (1975: 108) suggested, it is not assignable to the present species. Guzmán 6830 has a distinct bulb; a thin, but distinct pileipellis; and remnants of volval material containing plentiful filamentous, undifferentiated hyphae and plentiful inflated cells (subglobose to ovoid to ellipsoid to clavate, terminal, singly or in chains); it might be assignable to Amanita subsection Solitariae, but not to subsection Vittadiniae. Unfortunately, the specimen in question has lamellae damaged sufficiently that the presence or absence of clamps could not be determined after examination of several sections. The present species was reported from Anjou by Mornand and Galand (1991) and from Vendée by Pacaud (1997). I have examined the three collections discussed by Mornand and Galand and found them not assignable to A. subcaligata. It seems probable to me that their material from Anjou represents A. codinae. At present, I have not been able to review any material from Vendée. | ||||||||||||||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita subcaligata |
| bottom links |
[ Keys & Checklists ] |
| name | Amanita subcaligata |
| bottom links |
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.