


| name | Amanita strobilaceovolvata |
| name status | nomen acceptum |
| author | Beeli |
| english name | "Pine Cone Volva Ringed Ringless Amanita" |
| intro |
The following description is based on Beeli (1935). |
| cap | The prominently umbonate cap of Amanita strobilaceovolvata is 60 - 120 mm wide, fleshy, conic-convex, glabrous, dark brown in the center and for the inner half of the cap's radius and otherwise yellow; it is dry, with a striate margin. The cap skin can be pealed off the underlying flesh. The cap flesh is firm and white—except for being yellowish below the cap skin. |
| gills | The gills are free, somewhat pointed at both ends, crowded, yellowish-white, 4 mm broad, thin, and fragile. There are two short gills for every seven gills extending from margin to stem. |
| stem | The hollow stem of this mushroom is 120 - 220 × 7 - 10 mm, cylindric, fibrillose, smooth or sub-smooth, easily separated from the cap, and yellowish white. The ring is membranous, white, superior, skirt-like, and fragile. The flesh is firm and white. The volva is saccate, pinkish white; the exterior surface is divided into lumpy or bulging regions so that it looks like a net bag full of fruit. The volva sac is connected only at the stem's very base as depicted in Madame Goossens' watercolor. The fleshy interior of the volva is orangish-brownish. |
| odor/taste | This mushroom is odorless, and the taste is slightly acrid. |
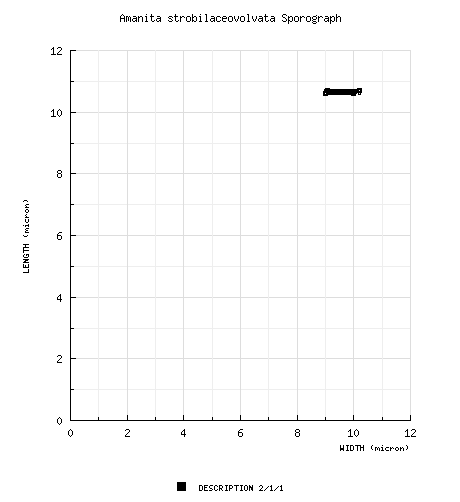
| spores | Based on the spore drawings of Gilbert (1941), the spores are 9.3 - 11.3 × 8.1 - 9.5 (-9.9) µm, subglobose to broadly ellipsoid and inamyloid. |
| discussion |
The present species was originally described from
the Democratic Republic of Congo where it is said
to occur singly in dry forest. Gilbert (1941) makes the case that Amanita fibrillosa Beeli was based upon an old specimen of Amanita strobilaceovolvata. However, Gilbert did not take into account several observations that do not support his proposal. These are discussed on the taxon page for A. fibrillosa. A number of African species are described as having a decorated exterior to the volva usually surface cracking producing darkened areolae, for example, Amanita zambiana Pegler & Piearce. However, this form of patchiness on the volval exterior is different from bumpy decoration of the volval sac in the present species and is known from volval surfaces of species belonging in both sect. Caesareae and sect. Amanita (e.g., A. pudica. Zhu L. Yang has described species of sect. Vaginatae with decorated volva from Yunnan Province, China. One such example is A. verrucosivolva Zhu L. Yang (1997). But in this species, while the exterior of the volval sac is divided into many regions, the regions do not bulge outward and are each decorated by a single, central wart. The reader may wish to refer to other species of section Vaginatae with an annulate stipe. Among these are Amanita madagascariensis (warted universal veil) and A. sp-Arora_01-555 (smooth universal veil). Sporograph comparisons are available on the technical tab of this page. Because this species is an annulate species of section Vaginatae, as are the other speices assigned to to the present stirps, it is assigned here despite lack of molecular evidence. Amanita strobilaceovolvata sensu Pegler and Shah-Sm. was demonstrated to be based on a mixture of distinct taxa by Tang et al. (2015).—R. E. Tulloss |
| brief editors | RET |
| name | Amanita strobilaceovolvata | ||||||||
| author | Beeli. 1935. Fl. Iconogr. Champ. Congo 1: 14, tab. 1 (fig. 4). | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Pine Cone Volva Ringed Ringless Amanita" | ||||||||
| synonyms |
≡Amanitopsis strobilaceovolvata (Beeli) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 75, tab. 6 (figs. 1-2). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 479212, 284116 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | BR (implicit) | ||||||||
| selected illustrations | E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl.: 225, tab. 8. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present taxon and from (Gilbert 1940 & 1941). | ||||||||
| pileus | from protolog: 60 - 120 mm wide, fuligineous on yellow ground (i.e. (per figure), fuligineous over disc and outward about one-half of radius and yellow for remainder of radius), conic-convex, umbonate, glabrous, dry; context fleshy, firm, white, [?except—ed.] yellowish under pileipellis; margin striate (ca. 0.6R per figure); universal veil absent; pileipellis separable. | ||||||||
| lamellae | from protolog: free, crowded, white with yellowish tint, 4 mm broad, subacute at both ends, thin, fragile and breaking, 7 per cm (?at pileus circumference); lamellulae 2 per cm (?at pileus circumference). | ||||||||
| stipe | from protolog: 120 - 220 × 7 - 10 mm, yellowish white, cylindric, smooth or subsmooth, easily separable from pileus, totally elongating per figure; context hollow, firm, white; partial veil superior, membranous, white, pendent, fragile; universal veil as saccate volva, with exterior divided into bulging regions [like net bag of fruit per figure—ed.], membranous, thick, with pallid to reddish brown tinted exterior and orangish-brownish in interior. | ||||||||
| odor/taste | from protolog: Odor lacking. Taste very slightly acrid. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | from protolog: at surface filamentous hyphae 2 μm wide, subhyaline, interwoven "parallelly." | ||||||||
| pileus context | not described. | ||||||||
| lamella trama | not described. | ||||||||
| subhymenium | not described. | ||||||||
| basidia | not described. | ||||||||
| universal veil | not described. | ||||||||
| stipe context | from protolog: at surface filamentous hyphae 4 - 5 μm wide, parallel. | ||||||||
| partial veil | not described. | ||||||||
| lamella edge tissue | not described. | ||||||||
| basidiospores |
from protolog: 6 μm wide, hyaline, smooth, inamyloid, "globose"; apiculus present (per figure); contents not described; white in deposit. Gilbert (1940): [2/1/1] 10.6 - 10.7 × 9.0 - 10.2 μm, (Q = 1.06 - 1.18), hyaline, smooth, inamyloid, subglobose to broadly ellipsoid, at least sometimes adaxially flattened (per figure), inamyloid; apiculus sublateral and cylindric to truncate-conic (all per figure); contents not described; white in deposit. [Note: Spore measurements are taken from two drawings of (Gilbert 1940: tab. VI (fig. 1)) that are most likely to be presented in lateral view.—ed.] | ||||||||
| ecology | from protolog: Solitary. Terrestrial in dry forest. | ||||||||
| material examined | from protolog: CONGO, DEMOCRATIC REPUBLIC OF: PROV. EQUATEUR—Territoire Lisala - Binga [2°23'41" N/ 20°25'25" E, 361 m], vi.1931 M. Goossens-Fontana 928 (holotype, BR, watercolor). | ||||||||
| discussion |
For a discussion of why Gilbert's proposal
(1941) to
treat A. fibrillosa as a taxnomic synonym of
the present species appears not to be well-supported,
see the discussion on the technical tab of
A.
fibrillosa. Amanita strobilaceovolvata sensu Pegler and Shah-Sm. was demonstrated to be based on a misture of distinct taxa by Tang et al. (2015). However, Tang et al. fail to demonstrate that the specimen they examined morphologically satisfies the original description of the present species or the partial type review of Gilbert (1940-1941). For example, the cap color is not a match for that of the present species as they claim; and Gilbert's spore data (which RET has often found to be reliable so far as it goes) falls entirely outside of the size-shape ranges reported by Tang et al. The sequence they propose for the species comes from a specimen that is not mentioned at all in terms of morphological revision. It seems they have not resolved the concept of A. strobilaceovolvata. The following figures compare the known spore data of the present species with sporographs of other taxa of section Vaginatae that have an annulate stipe—A. madagascariensis (warted volval surface) and A. sp-Arora_01-555 (smooth volval surface): | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita strobilaceovolvata |
| bottom links |
[ Keys & Checklists ] [ Subsaharan List ] [ Draft description of, & key to, sect. Caesareae ] |
| name | Amanita strobilaceovolvata |
| bottom links |
[ Keys & Checklists ] [ Subsaharan List ] [ Draft description of, & key to, sect. Caesareae ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.