


| name | Amanita silvicola |
| name status | nomen acceptum |
| author | Kauffman |
| english name | "Kauffman's Forest Amanita" |
| images |
 1. Amanita silvicola - C. Bas drawing  2. Amanita silvicola, Jackson Co., Oregon, U.S.A. ") 3. Amanita silvicola, Gifford Pinchot, Nat. For., Multnomah Co., Washington, U.S.A. (RET 456-4) |
| intro |
The following is largely based on the work of Bas (1969). |
| cap |
The cap of Amanita silvicola is 50 - 120 mm wide, convex, with a slightly flattened center to plano-convex or flat, with a nonsulcate, appendiculate margin long remaining incurved. The cap is at first completely covered by a rather thin, soft, white to sordid, floccose, continuous layer of volva, later often with scattered, large to small, floccose to felted, irregular patches of volva on a subviscid, white cap skin. |
| gills |
The gills are crowded, free to narrowly adnate, rather narrow to moderately broad, and white with a flocculose edge. The short gills are rounded-subtruncate to attenuate. |
| stem |
The stem is 60 - 100 × 15 - 25 mm, subcylindrical, solid, white, floccose, exannulate or with evanescent floccose annular zone, with floccose to felted remnants of volva on the outside of the subabrupt to abrupt, often marginate, rather top-shaped bulb (up to 30 mm wide). The bulb may lose its abrupt character with age and become closer to clavate without a distinct margin. |
| odor/taste |
Throughout the fruiting body, the flesh is white and soft. Odor and taste are reportedly indistinct. |
| spores | The spores measure (6.1-) 7.2 - 10.0 (-12.5) × (4.0-) 4.2 - 6.0 (-8.8) µm and are amyloid and ellipsoid to elongate or (infrequently) cylindric. Clamps are absent at bases of basidia. |
| discussion |
The present species was originally described from Oregon, U.S.A. Within the U.S.A., the species is also known from the states of California and Washington. I believe its range may extend across the Sierra Nevada and include at least part of Montana. In Canada, the species is also known from the Prov. British Colombia. The type collection was made in a dense forest of Hemlock (Tsuga) and Thuja. Dr. Bas (1969) placed Amanita silvicola as the sole taxon in his stirps Silvicola.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita silvicola | ||||||||||||||||||||||||||||||||||||
| author | Kauffman. 1926. Pap. Michigan Acad. Sci. 5: 123, pl. 4. | ||||||||||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||||||||||
| english name | "Kauffman's Forest Amanita" | ||||||||||||||||||||||||||||||||||||
| synonyms |
≡Aspidella silvicola (Kauffman) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 79, tab. 55 (fig. 4). The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||||||||||||||||||
| etymology | sylva or silva ("forest" or "wood" or "woodland") + -icola ("inhabitant of") | ||||||||||||||||||||||||||||||||||||
| MycoBank nos. | 278165, 284331 | ||||||||||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||||||||||
| holotypes | MICH | ||||||||||||||||||||||||||||||||||||
| revisions |
Bas. 1969. Persoonia 5: 526, figs. 314-316. Tulloss and Lindgren, here | ||||||||||||||||||||||||||||||||||||
| selected illustrations | E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27, suppl.: tab. 67. | ||||||||||||||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following is based on material derived from the protolog (Bas 1969) and original research of R. E. Tulloss and J. E. Lindgren. | ||||||||||||||||||||||||||||||||||||
| pileus | 50 - 120 mm wide, white, sometimes becoming slightly sordid, sometimes faintly pinkish in scattered areas, convex with slightly flattened disc to plano-convex or plane, dull to subshiny when dry; context white, quite firm when young, 12.5+ mm thick over stipe, thinning evenly to margin; margin nonstriate, appendiculate with quickly drying fibrillose material, incurved at first, remaining decurved at maturity, subviscid; universal veil as continuous floccose layer at first, thin, soft, white to sordid (similar to change of pileus surface), later breaking up into scattered irregular patches, variously sized, floccose to subfelted to felted, sometimes quite thin suggesting hoar frost, sometimes minimal to absent near margin. | ||||||||||||||||||||||||||||||||||||
| lamellae | free to narrowly adnate with decurrent tooth, often with decurrent line on stipe apex (marked in young material), crowded, white in mass, very pale cream in side view, not changing when cut or bruised, 7+ mm broad, broadest at 0.6 - 0.75R, projecting below pileus margin at maturity, with flocculose edge; lamellulae truncate to subtrucate to rounded truncate to attenuate, unevenly distributed, plentiful, of diverse lengths. | ||||||||||||||||||||||||||||||||||||
| stipe | 47 - 100 × 12 - 25 mm, white, becoming faintly yellow or sordid after handling, cylindric to subcylindric, flairing just at very apex, longitudinally striatulate, with some raised filaments darkening from handling, quite flocculent for top 15+ mm of stipe at first (showing marks of lamella edges), floccose; bulb marginate, turbinate to subnapiform, becoming immarginate and ellipsoid to clavate to slenderly clavate, 30 - 32+ × 24 - 30 mm wide, with rounded and rarely rooting base; context white to off-white, soft, solid or with stuffed central cylinder 8+ mm wide, with stuffing material of white longitudinally oriented fibers; partial veil absent or as an evanescent floccose annular zone; universal veil sometimes as floccose to felted fragments forming fluffy or somewhat firm rim at top or margin of bulb. | ||||||||||||||||||||||||||||||||||||
| odor/taste | Odor at first not distinct or faintly flower-like or like very old peaches, finally marked "as of very alkaline urine or potash." Taste not recorded. | ||||||||||||||||||||||||||||||||||||
| macrochemical tests |
Spot test for tyrosinase (paracresol) - negative throughout basidiome. Spot test for laccase (syringaldazine) - negative throughout basidiome or positive in a small region near surface of upper part of bulb. Test voucher: Lindgren 91-38. Results according to Breckon (1968)—on exposed context of the lower stipe and bulb unless otherwise noted: FeSO4 - pale peach color; 3% KOH - negative; Melzer’s reagent - negative; phenol - negative; phenolaniline - negative; tincture of guaiac - negative. | ||||||||||||||||||||||||||||||||||||
| basidia | Bas (1969): 40 - 55 × 8 - 10 μm, mostly 4-, occasionally, some 2-sterigmate; clamps lacking. | ||||||||||||||||||||||||||||||||||||
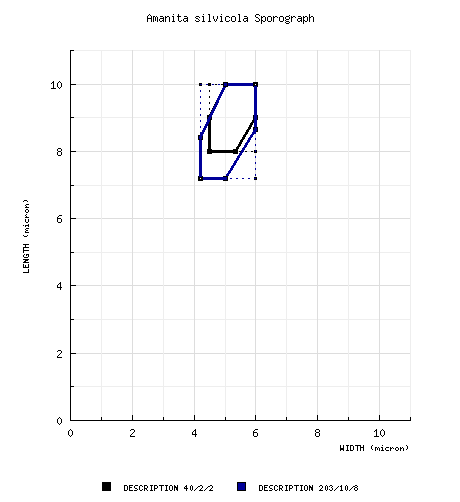
| basidiospores |
Bas (1969):
[40/2/2] 8.0 - 10.0 (-11.5) × 4.5 - 6 μm, (Q = 1.50 -
2.0; Q = 1.65 - 1.85), colorless, hyaline,
smooth, thin-walled, amyloid, ellipsoid to elongate,
sometimes ovoid; apiculus often rather strongly
projecting; contents subgranular, refractive;
color in deposit not recorded. RET: [203/10/8] (6.1-) 7.2 - 10.0 (-12.5) × (4.0-) 4.2 - 6.0 (-8.8) µm, (L = (7.5-) 8.0 - 9.2 µm; L' = 8.5 µm; W = 4.5 - 5.4 (-6.0) µm; W' = 5.1 µm; Q = (1.31-) 1.44 - 2.0 (-2.18) µm; Q = (1.52-) 1.62 - 1.83 µm; Q' = 1.69 µm), hyaline, colorless, smooth, thin-walled, amyloid, ellipsoid to elongate to cylindric; apiculus sublateral, cylindric, often prominent; contents multiguttulate; white in deposit. | ||||||||||||||||||||||||||||||||||||
| ecology |
Bas (1969): [Oregon:] "...type collected in dense Tsuga-Thuja forest with underbrush." Solitary to scattered. California: Under Pinus. Washington: Under Pseudotsuga menziesii. Oregon: Under leaves in Quercus thicket or under mixed conifers or in Tsuga-Thuja forest with underbrush. [NOTE: See also Hotson (1936) and Bandoni & Szczawinski (1964).] | ||||||||||||||||||||||||||||||||||||
| material examined |
Bas (1969): U.S.A.: OREGON—Clackamas Co. - Mt. Hood, 30.ix.1922 C. H. Kauffman s.n. (holotype, MICH).
WASHINGTON—Clallum Co. - Joyce, 28.x.1935
A. H. Smith 3398 (MICH; L; photograph in [Gilbert
1941:
pl. 67]). RET: U.S.A.: CALIFORNIA—Alameda Co. - Livermore, grounds of Lawrence Livermore Laboratories, 27.i.2003 Tom Humphrey s.n. (RET 366-10). Santa Cruz Co. - ca. Santa Cruz, 15.i.2003 FFSC member s.n. [Tulloss 1-15-03-C] (RET 366-1), 4.xii.2004 John Brown s.n. (??; RET 382-4). OREGON—Benton Co. - ca. Corvallis, xi.1921 Carl Clawson Epling 333 (MICH as "A. chlorinosma sensu Kauffman" per Zeller). Jackson Co. - Rte. 230 milemarker 6, 19.x.2010 Ron Pastorino 10-19-10A [mushroomobserver.org # 56201] (RET 454-2). Tillamook Co. - Tillamook St. For., ca. Brown Camp, 31.x.2010 Sava Kristić s.n. [mushroomobserver.org # 57708 p.p.] (RET 589-7). WASHINGTON—Island Co. - Whidbey Isl., 2.4 km N of Columbia Beach, 28.x.1962 D. Grund 712 (ACAD 10682). Multnomah Co. - SE GPNF, 10.x.2010 Sava Krstić s.n. [mushroomobserver.org # 55419] (RET 456-4). King Co. - North Bend, North Cascades Nat. Pk. [ca. 47.488° N/ 121.705° S, 311 m], 14.x.2013 unkn. coll. s.n. [mushroomobserver #148538] (RET 594-9, nrITS & nrLSU seq'd.). Thurston Co. - ca. Foster, S of Olympia, 8.xi.1958 unkn. coll. Is-766 (WTU). Unkn. Co. - in Olympic Mtns., 12.x.1915 C. H. Kauffman s.n. (MICH as "A. solitaria"). [NOTE: See Thiers 43984 (SFSU).] | ||||||||||||||||||||||||||||||||||||
| discussion |
t.b.d. RET 589-7 is immature. [Note: We need to get some very young material of A. silvicola to check to see if there is a thin, hyphal outerlayer to the volva.—RET] | ||||||||||||||||||||||||||||||||||||
| citations | —R. E. Tulloss and J. E. Lindgren | ||||||||||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita silvicola |
| name status | nomen acceptum |
| author | Kauffman |
| english name | "Kauffman's Forest Amanita" |
| images |
1. Amanita silvicola - C. Bas drawing 2. Amanita silvicola, Jackson Co., Oregon, U.S.A. 3. Amanita silvicola, Gifford Pinchot, Nat. For., Multnomah Co., Washington, U.S.A. (RET 456-4) |
| photo |
Ron Pastorino - (2) Jackson County, Oregon, U.S.A. [mushroomobserver.org/56201]. Sava Krstić - (3) Gifford Pinchot National Forest, Multnomah County, Washington, U.S.A. (RET 456-4) [mushroomobserver.org/55419]. |
| drawing | Dr. Cornelis Bas (1969) - (1) Pacific Northwest of the U.S.A., courtesy of Persoonia, Leiden, the Netherlands. |
| name | Amanita silvicola |
| bottom links |
[ Keys & Checklists ] |
| name | Amanita silvicola |
| bottom links |
[ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.