


| name | Amanita populiphila |
| name status | nomen acceptum |
| author | Tulloss & E. Moses |
| english name | "Poplar-loving Ringless Amanita" |
| images |
 1. Amanita populiphila, Colin Nesblett Wildlife Area, Colfax Co., New Mexico, U.S.A.  2. Amanita populiphila, Colin Nesblett Wildlife Area, Colfax Co., New Mexico, U.S.A.  3. Amanita populiphila, Turtle Crk. St. Pk., Pottawatomie Co., Kansas, U.S.A.  4. Amanita populiphila, Colin Nesblett Wildlife Area, Colfax Co., New Mexico, U.S.A.  5. Amanita populiphila, after heavy rain, Nebraska, U.S.A. |
| intro | This species was originally described by Tulloss and E. Moses (1995). |
| cap | Amanita populiphila has a cap 30 - 120 (-135) mm wide that is white to off-white to pale tan or pale cream at first and becomes pale yellowish to straw-color to tan to brown with age and handling. It is broadly campanulate at first, then convex and broadly umbonate, and finally plano-convex in age with its margin uplifted. The flesh is 5 - 12.5 mm thick over the stem and is pure white, unchanging with cutting or bruising. The cap margin is striate (about 15% of the radius at first, becoming 20% to 40% of the radius).The volva may be absent or present as warts, small patches, or a single patch with extensive cracking on its surface. The volva is white at first and tends to change in color as the cap does (even occasionally becoming significantly darker than the cap); it is fragile, and easily removed. |
| gills | The gills of this species are free or narrowly attached, sometimes leaving very pallid orange vertical lines on the upper stipe; they are close to crowded and off-white to cream with pale orangish or pinkish tints in mass. In side view, the gills are white to various shades of cream and do not change when cut or bruised. Just after drying they are often a pale shade of pink or orange. They are 3 - 7.5 mm wide and have flocculence on the edge that ranges from concolorous to pale orangish white. Short gills are truncate to excavate-truncate, in several ranks, unevenly distributed, and common to plentiful. |
| stem | The stem of A. populiphila is 63 - 175 (-220) × (5-) 7 - 20 mm, white (sometimes with pale watery streaks), sometimes becoming pale watery brown after handling (but often not changing), and exannulate. The flesh is white to off-white and usually unchanging when cut or bruised (concolorous in tunnels made by insect larvae). There is a hollow central cylinder 2.5 - 14 mm wide. The volva is white (with color changes as on the cap) and more cup-shaped than sac-shaped. It is weakly structured and may break up into warts or patches on the lower stipe or simply come off the stem entirely during collecting (like a thimble) unless carefully excavated with the stem base. |
| odor/taste |
Amanita populiphila has a mild or insignificant odor at first that may become pungent in older specimens. The Moses family has eaten and enjoyed this species, they say the taste is mild and "pleasantly fungoid" except for the tasteless volva; and, in their preparation, the stem was "pleasantly crispy." |
| spores | The spores of this species measure (7.0-) 9.2 - 12.5 (-21) × (6.0-) 8.2 - 11.2 (-15.8) µm and are predominantly subglobose (occasionally globose or broadly ellipsoid) and inamyloid. Clamps are not found at the bases of basidia. |
| discussion | Amanita populiphila wasm first found under Cottonwood and Aspen (Populus) in the central and western USA—from Michigan and Kansas westward to Idaho, Wyoming, Colorado, and New Mexico. In 2008, a collection of this species by Raymond McNeil (again found under Populus) was sent to me by Renée Lebeuf from Prov. Québec, Canada. This is the most easterly known site for the species at present. This species was distinguished in detail from A. nivalis Grev. and A. vaginata var. alba Gillet in its original description.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita populiphila | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| author | Tulloss & E. Moses. 1995. Mycotaxon 53: 455, figs. 1-4. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| english name | "Poplar-loving Ringless Amanita" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| etymology | populus + -philus; hence, Populus-loving, because of its association with species of the genus Populus | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MycoBank nos. | 363198 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| holotypes | NY; isotype in RET | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| revisions | Tulloss, here. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon is based upon original research by R. E. Tulloss. The protolog of A. populiphila is available on this site. (here) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileus | protolog: 30 - 120 (-135) mm wide, white to off-white to Tilleul Buff (7.5YR 8.4/2.0), very pale tan (slightly darker over disc) or pale cream (with orange-brown tint over disc), often becoming yellowish (e.g., more cream than 5A2 with margin remaining nearly white) to straw-color to tan to brown with age and handling, tacky to subviscid to viscid (Smith 50210), shiny to dull, broadly campanulate, then convex and broadly umbonate, then plano-convex, in age with margin uplifted; context pure white, unchanging when bruised or cut, 5 - 12.5 mm thick at stipe, thinning evenly for up to four-fifths of radius, then membranous to margin; margin rather short striate at least at first (0.15±R), eventually with longer striation (0.2 - 0.4R) particularly in dry weather, frequently becoming rimose, nonappendiculate; universal veil absent or in often rather thick warts or small patches or as large areolate patch (Smith 84263), submembranous, fragile, detersile, off-white to avellaneous at first, unchanging or becoming yellowish brown or yellowish buff to pale brown to grayish brown to "very dark" (Trueblood 5472) in age or through drying or handling. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamellae | protolog: free to adnexed, sometimes with long decurrent very pale orangish white lines on upper stipe, close to crowded, off-white to pinkish white to very pale orangish or pinkish cream in mass, cream to pale cream to sordid pale cream to white in side view, unchanging when cut or bruised, pale pinkish or salmon pink (Trueblood 5472) or pale orange to pale yellow-orange when just dried, 3 - 7.5 mm broad, with concolorous to very pale orangish white flocculence on edge; lamellulae truncate to excavate-truncate, in several ranks, unevenly distributed, common to plentiful. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| stipe | protolog: 63 - 175 (-220) × (5-) 7 - 20 mm, white, sometimes with pale watery streaks, not discoloring or becoming watery pale brown over time or from handling, narrowing upward to subcylindric, flaring slightly or not at all at apex, decorated in upper half with very pale orangish white flocculence, finely striatulate; context white to off-white (with watery streaks), usually unchanging when cut or bruised, sometimes sordid in lower stipe or pale tan in universal veil below stipe base, concolorous in larva tunnels, stuffed in whole or part with white cottony fibrils, becoming hollow, with central cylinder 2.5 - 14 mm wide; exannulate; universal veil as "thimble-like" cupulate volva, submembranous to membranous, thin to 3.5 mm or more thick, white, changing with age and drying as on pileus or taking on orange-red or brown spots in lowest portion, easily detached from stipe in mature specimens, frequently also as warts (separate or confluent) unevenly distributed along stipe or in rings around stipe and having form like that of warts on pileus. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| odor/taste | protolog: Odor none (Smith 50210) or mild and pleasantly fungoid or pungent (Smith 84263) in older specimen. Taste mild and pleasantly fungoid in all parts except universal veil (tasteless). EDIBLE (with caution) and good with exception of unavoidably gritty universal veil [per E. Moses—ed.]. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| macrochemical tests |
protolog: Paracresol spot test for tyrosinase - for freshly collected material, rapidly positive throughout basidiome in specimens of all stages of development; in material collected 26 hours before testing, reaction took 5 min. Syringaldazine spot test for laccase - negative throughout basidiome with exception of exterior surface of undried portions of universal veil at stipe base and there rather rapidly and strongly positive (reddish lavender instantly or in less than 1 min., deepening to near ruby red). 95% ethanol (control for syringaldazine test) - negative in all parts. KOH - negative on pileus and lamellae. NH4OH - very pale yellow on context throughout basidiome. Test vouchers: holotype and Moses 1. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileipellis | protolog: with rather extensively gelatinized suprapellis colorless and 15 - 45 µm thick, with ungelatinized subpellis yellowish and 135 - 200 µm thick; filamentous, undifferentiated hyphae 1.8 - 6.5 µm wide, branching, densely packed, subradially arranged, sometimes with slightly thickened walls; vascular hyphae 2.2 - 17.0 µm wide, common to plentiful, branching, sometimes tightly coiling in part, best seen in scalp. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileus context | protolog: filamentous, undifferentiated hyphae 2.8 - 14.8 µm wide, plentiful to dominant, frequently branching, loosely interwoven, only infrequently in fascicles, with walls thin or up to 0.8 µm thick, often constricted at septa, with occasional slightly inflated intercalary segments up to 15.0 µm wide; acrophysalides plentiful, elongate to narrowly clavate, up to 121 × 34 µm, with walls thin or up to 0.8 µm thick; vascular hyphae 3.5 - 15.0 µm wide, common, sinuous, occasionally branching; clamps not observed. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamella trama | protolog: bilateral, with shallow angle of divergence; wcs = (50-) 60 - 85 µm when rehydrated well; subhymenial base containing numerous intercalary thin-walled inflated cells [clavate or obclavate to ellipsoid (up to 48 × 21 µm) to fusiform or elongate or allantoid (up to 85 × 20 µm) to (rarely) globose (e.g., 23 × 22 µm)]; filamentous, undifferentiated hyphae 1.5 - 11.0 µm wide, branching, with plentiful intercalary partially inflated to subfusiform to fusiform to broadly fusiform to clavate cells in central stratum (up to 52 × 17.5 µm or larger, becoming disoriented rather easily by sectioning); divergent, terminal, inflated cells not observed; vascular hyphae 2.0 - 4.8 µm wide, sinuous, observed in only one mount (Solheim 5259)—rare. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| subhymenium | protolog: wst-near = 40 - 75 µm when rehydrated well; wst-far = 70 - 100 µm when rehydrated well; frequently branching and rather tangled structure of short uninflated or partially inflated hyphal segments and small inflated cells, with basidia arising mostly from uninflated to partially inflated hyphal segments or from uninflated to slightly inflated branched elements, with basidia arising occasionally from small inflated cells; clamps infrequent to rare. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| basidia | protolog: 36 - 83 × 7.8 - 17.8 µm, thin-walled, 4- or occasionally 2-sterigmate, with sterigmata up to 8.0 × 5.0 µm; clamps and proliferated clamps scattered (most often found near pileus margin), infrequently locally common. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| universal veil | protolog: On pileus: as on stipe base except exterior surface layer sometimes missing, often with much of remainder partially gelatinized; filamentous, undifferentiated hyphae in narrower fascicles than at stipe base and with diameter up to 16.2 µm, with tip cells often somewhat inflated; inflated cells up to 150± × 42 µm (almost always less than 65 × 40 µm), common to plentiful, often badly collapsed. At base of stipe, exterior surface: filamentous, undifferentiated hyphae 1.8 - 8.8 µm wide, partially gelatinized to gelatinized, sometimes with walls slightly thickened, almost always in thick criss-crossing fascicles, with occasional slightly inflated terminal segment (up to 10.8 µm wide); vascular hyphae not observed. At base of stipe, interior: filamentous, undifferentiated hyphae 1.8 - 14.5 µm wide, frequently branching, dominating, almost always in broad fascicles, with walls up to 1.0 µm thick, with some branches out of fascicles loosely coiling, occasionally having yellowish subrefractive walls; inflated cells unevenly distributed, occasionally in loose clusters (rather commonly in Trueblood 5472), terminal, sometimes dissociated, hyaline, colorless, with walls thin or up to 1.0 µm thick, pyriform to subpyriform to broadly clavate to clavate to elongate-constricted to subfusiform to ellipsoid to ovoid to subglobose, up to 101 × 68 µm (almost always not more than 60 × 40 µm); vascular hyphae not observed; clamps not observed. At base of stipe, inner surface: filamentous, undifferentiated hyphae 1.8 - 10.0 µm wide, branching, in densely packed thin layer, longitudinally oriented, partially to extensively gelatinized, fasciculate, often with thickened walls; vascular hyphae 5.0 - 8.5 µm wide, in scattered fragments, partially gelatinized. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| stipe context | protolog: longitudinally acrophysalidic; filamentous, undifferentiated hyphae 2.8 - 9.1 µm wide, with thin or slightly thickened walls, very frequently branching, often in rather robust fascicles, plentiful to dominant; acrophysalides plentiful, up to 264 × 42 µm, with walls thin or up to 0.5 µm thick; vascular hyphae 4.5 - 11.2 µm wide, sinuous, scattered, locally common; clamps not observed. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| partial veil | absent. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamella edge tissue | not described in protolog. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| anatomical figures |
, elements of hymenium, subhymenium, and subhymenial base.") 1. Amanita populiphila (holotype), elements of hymenium, subhymenium, and subhymenial base. , elements of internal layer of universal veil from stipe base.") 2. Amanita populiphila (holotype), elements of internal layer of universal veil from stipe base. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
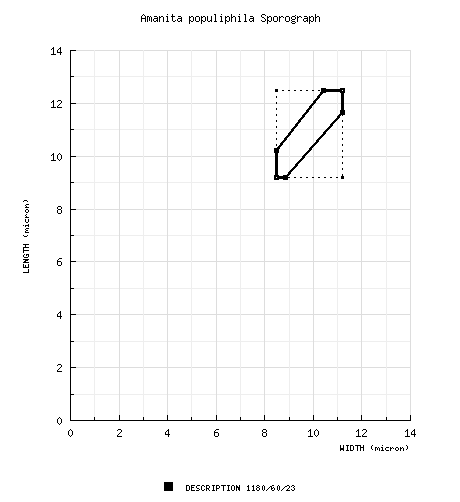
| basidiospores | composite from all material revised: [1180/60/23] (7.0-) 9.2 - 12.5 (-21) × (6.0-) 8.5 - 11.2 (-15.8) µm, (L = (9.8-) 10.1 - 11.9 (-12.2) µm; L’ = 10.9 µm; W = (9.0-) 9.1 - 10.7 (-11.1) µm; W’ = 9.8 µm; Q = (1.0-) 1.04 - 1.20 (-1.61); Q = (1.06-) 1.08 - 1.15 (-1.19); Q’ = 1.11), hyaline, colorless, smooth, thin-walled, inamyloid, subglobose, occasionally globose, occasionally broadly ellipsoid, infrequently lachrimiform or langeniform, very rarely ellipsoid, often adaxially flattened, often expanded at one end; apiculus sublateral to lateral, small, truncate-conic to cylindric; contents monoguttulate; white in deposit. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ecology |
Scattered to gregarious to
"occurring in hundreds." Québec, Canada:
Under Populus. U.S.A.: Colorado: At
1700± m elev. Under
Populus alba and P. sargentii in low
grassy area. Idaho: At
1900± m elev. Under Populus
tremuloides with (Solheim 5259) or without
(Trueblood 5472) nearby conifers.
Pottawatomie Co., Kansas: At 313-315 m elev.
August, 1990: occurring in hundreds (possibly with
several mycelia each producing 25 - 150 basidiomes)
covering several acres in mowed field (within flood
plain) under P. deltoides; with similar
large fruiting in September, 1989 (no voucher); with
May, 1991 fruiting producing no more than 50
basidiomes; with no fruiting in August, 1991; in
1992 (with plentiful rainfall and cool weather)
fruitings starting in mid-May, with close to 2000
basidiomes present simultaneously (probably from
twenty or more separate mycelia) by early June; with
fruiting reduced to several hundred fresh basidiomes
in third week of month; with fruiting continuing
with reduction in number and size of basidiomes
until only twenty found on 27 June. [Between
1 July and 25 July 1992, area had 28.8 cm of
rain.] 25 July 1992, 200 basidiomes counted
(most smaller than usual); 29 July, 1992, about
100 fresh basidiomes in main fruiting area with
scattered specimens in two other nearby
sites. [Temperatures began to rise in late
July, 1992 and became very hot in first week of
August.] 2 August, 1992, in continuing rain,
about 200 basidiomes found in several sites.
8 August, 1992, about 150 basidiomes (most smaller
than usual) total for all sites. [During
mid-August, weather remained hot and became
dry.] 18 August, 1992, 12 basidiomes in main
area of occurrence, with 3 in distant site and these
with darker than usual disc; 22 August, 1992: no
fruiting bodies seen. [July 1992 rainfall
(33.6 cm) was third highest recorded for that month
for eastern Kansas region in Twentieth
Century.] Michigan: On moss at edge of bog
(Smith 50210). Minnesota: In dark soil of old
lake bottom, under P. tremuloides or in
sparsely wooded area containing P.
tremuloides, Juniperus, and plentiful
young buckthorn (Rhamnus) scrub. New
Mexico: In sod of pasture with Populus sp.
[?fremontii] nearby (Barrows 1090) or in
dark wet loam of riparian habitat at
2410± m elev. with P. tremuloides,
Abies, and Acer negundo among low
cover including "cow parsnips," Rosa, and
Spirea (Tulloss 8-15-92-A). Wyoming:
In grassy area, under P.
tremuloides. [Note: Previous citation of Minnesota habitat did not refer to populiphila. See A. sp-MN03. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| material examined |
CANADA:
QUÉBEC—Région Montérégie - Châteauguay,
1.vii.2008 Raymond McNeil 86 (RET 447-1, nrITS &
nrLSU seq'd.).
U.S.A.:
COLORADO—Arapaho Co. - Littleton, Ketring
Pk., N of Ketring Lk. [39°36’31.2” N/ 104°59’58.63” W,
1700± m], 18.viii.1997 J. Justice, E.
Jacobson, M. Shaw & R. E. Tulloss 8-18-97-A
(DBG ??; RET 266-9,
nrITS & nrLSU seq'd.).
IDAHO—Fremont Co. -
Targhee Nat. For., 6.6 km W of Idaho/Wyoming state
line, 2.viii.1957 W. G. Solheim 5259 (paratype,
MICH). Owyhee Co. - Vulcan Crk., 7.vii.1973
Ellen & Ted Trueblood 5472 (paratype,
MICH).
KANSAS—Ellsworth Co. -
NE of Ellsworth, SW of Westfall, W of St. Rte. 156,
E-bound rest stop on I-70, ca. mile 224, 10.ix.2008
R. E. Tulloss 9-10-08-A (RET 423-2, nrITS & nrLSU
seq'd.). Pottawatomie Co. - N of Manhattan,
Tuttle Creek St. Pk., River Pond Area [39°15'05" N/
96°35'30" W, 314 m], 11-13.viii.1990 W. R. & E.
Moses s.n. [Tulloss 8-11/13-90-EM1] (paratype, RET
035-8, nrITS seq'd.), 25.viii.1990 W. R. & E. Moses
s.n. [Tulloss 8-25-90-EM1] (paratype, RET 036-1, nrITS
seq'd.), 25.v.1991 W. R. & E. Moses s.n. [Tulloss
5-25-91-EM1] (paratype, RET 047-3), 15.v.1992 W. R.
& E. Moses s.n. [Tulloss 5-15-92-EM1] (holotype,
NY; isotype, RET 119-1), 22.v.1992 W. R. & E. Moses
s.n. [Tulloss 5-22-92-EM1] (paratype, RET 60-4),
25.v.1992 W. R. & E. Moses s.n. [Tulloss
5-25-92-EM1] (paratype, L; paratype, RET 060-7),
30.v.1992 W. R. & E. Moses s.n. [Tulloss
5-30-92-EM1] (paratype, RET 103-6, nrITS &
nrLSU seq'd.), 5.vi.1992 W. R. & E. Moses 1
(paratype, NY), 2 (paratype, RET 103-3, nrITS
& nrLSU seq'd.), 3 (paratype, RET 103-4,
nrITS & nrLSU seq'd.), 4 (paratype, RET 103-5,
nrITS & nrLSU seq'd.). Rooks Co. -
Stockton, 1.ix.1908 Elam Bartholomew s.n. (paratype,
FH as "A. nivalis"). Unkn. Co. -
unkn. loc., 18.vi.1967 Harbaugh 67-6-18-1 (paratype,
MICH).
MICHIGAN—Luce Co. - Tahquamenon St. Pk.,
22.viii.1965 Alexander H. Smith 50210 (paratype,
MICH). Oakland Co. - Highland Recreation
Area, 22.vii.1973 A. H. Smith 84263 (paratype,
MICH).
MONTANA—Gallatin Co. -
Bozeman, divide btwn. Bridger & Kelly Cyn.,
2.vii.1957 F. B. Cotner 570039 (paratype,
MICH).
NEBRASKA—Dawson Co. -
ca. 2 km E of Cozad, Cozad W-bound Rest Stop, I-80,
2.ix.2008 R. E. Tulloss 9-2-08-A
(RET 423-6).
NEW MEXICO—Colfax Co. -
Colin Nesblett Wildlife Area, Tolby Creek
[2410± m], 15.viii.1992 NAMA ‘92 foray
participant s.n. [Tulloss 8-15-92-A] (paratype,
RET 068-7, nrITS seq'd.). Rio Arriba Co. -
El Rito, viii.1959 Charles Barrows 1090 (paratype,
MICH).
PENNSYLVANIA—Northampton Co. - Blue
Mountain, Game Lands 168, off Old Grade Rd.
[40.83025° N/ 75.35626° W, 271 m], 4.viii.2012 David
Wasilewski s.n. [Tulloss 8-4-12-F]
(RET 506-5, nrITS & nrLSU seq'd.).
WYOMING—Teton Co. - Grand Teton Nat. Pk.,
Reid Mtn., 26.viii.1983 Kent H. McKnight & Meinhard
Moser [McKnight 83082610] (paratype, BPI as "A.
alba"). [Note: A nonconformant paratype from Minnesota has been removed. See A. sp-MN03.] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| discussion |
t.b.d.Dr. L. V. Kudzma has successfully derived nrITS from the isotype, eight paratypes, and five recent collections of A. populiphila. Five single nucleotide polymorphisms (SNPs)are detectable in the otherwise invariant sequences. The first character of the 5' end of nrITS is taken to be the character immediately following the "GGATCATTA" 3' terminal motif of the 18S gene and is designated as character 1. If we treat populiphila as determined morphologically and having nrLSU sequences as described below. The nrITS sequences from twelve of the samples produce an alignment revealing 5 SNPs with positions as follows:The protolog of A. populiphila is available on this site. (here)character 14 - G or CWith regard to the nrLSU gene, Dr. Kudzma's nine sequences cumulatively cover the first 1455 characters where character 1 is determined to be the first character of the 5' terminal motif of the nrLSU gene. These sequences are inavariant with the exception of three SNPs and a one character indel (that might be a PCR artifact) as follows: | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.