


| name | Amanita heterochroma |
| name status | nomen acceptum |
| author | S. Curreli ex S. Curreli |
| english name | "Curreli's Variable Amanita" |
| synonyms |
≡Amanita variabilis S. Curreli nom. illeg.
non Amanita variabilis E.-J. Gilbert & Cleland |
| intro | The following is based on the original description of Amanita heterochroma by Curreli (2000) and the recent description of Neville and Poumarat (2004). |
| cap | The cap of A. heterochroma is 80 - 150 mm wide, fleshy, yellow-greenish tone when young, light brown out from the center, entirely brown at maturity, at first globose, sometimes with slight depression when fully expanded, viscid in humid weather, usually with a smooth margin. Marginal striations appear rarely in mature material, and then only faintly. The volva is present as plentiful warts that are ash-gray. The flesh is white and firm. |
| gills | The gills are free, rather crowded, narrow markedly toward the stem, at first white, then cream-yellowish ochraceous, with a finely floccose edge. The short gills are truncate and of diverse lengths. |
| stem | The stem is 100 - 180 × 15 - 25 mm, cylindric or slightly narrowing upward, smooth, white and smooth above the ring, ash gray becoming brown at maturity below the ring. The bulb is turnip-shaped, is more or less rooting, and has a point at the bottom. The ring is white, fragile, may be lost or remain only as a small fragment. The volva is friable, ash-gray, and forms numerous rings around the bottom of the stem and top of the bulb. The flesh is white and firm. |
| spores | The spores measure (8-) 9.5 - 15 (-22) x 6.7 - 8.2 µm and are ellipsoid to elongate and inamyloid. Clamps are rather numerous at bases of basidia. Neville and Poumarat (2004) report that the type of the present species is immature and lacks spores. Based on other material they report the following: (8.5-) 9 - 11 (-11.5) x (6-) 6.5 - 7.5 (-8) µm and are ellipsoid, rarely elongate and inamyloid. |
| discussion |
This species occurs in association with Eucalyptus camaldulensis and Cistus monspeliensis in sandy-gravelly soil. It is only known from Sardinia (Italy). Neville and Poumarat note that there is a slight possibility this species was introduced to Europe from Australia with Eucalyptus. On the other hand, the apparent close relationship to Amanita muscaria (L. : Fr.) Lam. suggests the present species may have evolved from a Eurasian ancestor. The reader should compare the present species with Amanita gioiosa S. Curreli ex S. Curreli since the two taxa appear in the same habitat. The best available description of the present species is that of Neville and Poumarat (2004).—R. E. Tulloss and L. Possiel |
| brief editors | RET |
| name | Amanita heterochroma | ||||||||
| author | ("heterochroa," "heterocroma") S. Curreli. 1992. Micol. Ital. 21(3): 15. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Curreli's Variable Amanita" | ||||||||
| synonyms |
≡Amanita variabilis S. Curreli nom. illeg. 1991. Micol. Ital. 20(1): 17 (unnumbered figs. on pp. 15-16). [Posterior homonym. ICBN §53.1] non Amanita variabilis E.-J. Gilbert & Cleland
≡Amanita muscaria var. heterochroma (S. Curreli) Contu. 2000. Boll. Gruppo Micol. G. Bressadola - Nuova Ser. 43(2): 78. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 467900, 355334, 467408 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes |
in herb. Gruppo Micologico Zonale San Gavino [Note: Immature per Neville and Poumarat (2002 ["2001"]. Bull. Trimestriel Soc. Mycol. France 117(4): 290.)] | ||||||||
| revisions | Neville and Poumarat. 2002 ["2001"]. Bull. Trimestriel Soc. Mycol. France 117(4): 289. | ||||||||
| selected illustrations | Curreli. 2000. Boll. Gruppo Micol. G. Bresadola 43(2): 91. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present species. | ||||||||
| basidiospores |
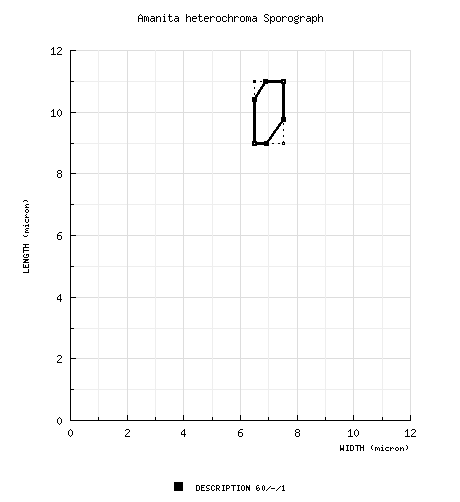
from protolog: 9.5 - 15 × 6.7 - 8.2 μm, (est. Q = 1.42 - 1.83), hyaline, smooth, inamyloid, ellipsoid to elongate; apiculus distinct; contents as single large guttule; cream-white in deposit. Neville & Poumarat (2004): [60/-/1] (8.5-) 9 - 11 (-11.5 × (6-) 6.5 - 7.5 (-8) μm, (L' = 9.9 μm; W' = 6.9 μm; Q = 1.30 - 1.60 (-1.70); Q 1.40 - 1.46; Q' = 1.43). | ||||||||
| ecology | from protolog: Under Eucaplyptus (introduced) and Cistus. | ||||||||
| material examined | from protolog: ITALY: SARDINIA—Prov. Cagliari - Sanluri, Sanluri Stato, 20.xii.1990 S. Curreli 260 (holotype, in herb. S. Curreli, San Gavino Monreale, Sardinia, Italy). | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.