


| name | Amanita codinae |
| name status | nomen acceptum |
| author | (R. Maire) Singer |
| english name | "Codina's Lepidella" |
| images |
.") 1. Amanita codinae, as illustrated by C. Bas (1969). |
| intro | This description was initially based on the review of this species by Bas (1969). Additional information is supplied from original research of RET. |
| cap | The cap of A. codinae is 50 - 90 (-130) mm wide, fleshy, convex, whitish to pale brownish, dry, appendiculate, with a nonsulcate margin. At the center, the cap is covered with adnate to detersile, pale brown to dark brown, small to rather coarse, felted-subfibrillose, subpyramidal to flat warts, towards the margin gradually passing into appressed, fibrillose scales. |
| gills | The gills are moderately crowded to crowded, adnexed to free, broad, white, and becoming cream to pale yellowish. The short gills are subtruncate to rounded or attenuate. |
| stem | The stem is 40 - 80 × 10 - 20 mm, cylindrical, solid, white, turning brownish when bruised, with mostly incomplete, circular zones of brownish to brown, fibrillose scales. |
| odor/taste | The odor of A. codinae is reported to be indistinct or faintly pleasant at first and to become slightly unpleasant. The taste is reported as mild. |
| spores | The spores measure (7.6-) 8.9 - 13.0 (-16.5) × (6.4-) 6.8 - 9.4 (-11.0) µm and are amyloid and ellipsoid to elongate. Clamps are abundant at bases of basidia. |
| discussion | This is a species of the Mediterranean region, known from Spain, southern France, and northern Morocco eastward. Reports from other parts of the world (e.g., the Americas) are apparently all based on incorrect determinations. Amanita codinae is similar to A. vittadinii (Moretti) Vitt. and other taxa in Bas' stirps Vittadinii. Like nearly all other members of subsection Vittadiniae, it is known to occur without woody plant symbionts.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita codinae | ||||||||||||||||||||||||||||
| author | (Maire) Bertault. 1955. Bull. Trimestriel Soc. Mycol. France 71: 28. | ||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||
| english name | "Codina's Lepidella" | ||||||||||||||||||||||||||||
| synonyms |
≡Amanita codinae (Maire) Singer nom. inval. 1951 ["1949"]. Lilloa 22: 388. [Not accepted by author in original publication. ICBN § 34.1(b)]
≡Lepidella codinae Maire. 1933. Mus. Barcin. Scient. Nat. Op. (Bot. 2) 15: 85, pl. 1.
≡Amanita vittadinii var. codinae (Maire) Veselý. 1934d. Atlas Champ. Eur. 4-5: 56.
≡Amanita vittadinii f. codinae (Maire) E.-J. Gilbert. 1941b. Notules Amanites: [3].
≡Armillaria codinae (Maire) Locq. 1952. Bull. Trimestriel Soc. Mycol. France 68: 167. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||||||||||
| etymology | genitive of Latinized name; hence, "of Codina" or "Codina's" | ||||||||||||||||||||||||||||
| MycoBank nos. | 292446, 261263, 281962, 292693 | ||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||
| holotypes | in herb. R. Maire => MPU | ||||||||||||||||||||||||||||
| revisions | Bas. 1969. Persoonia 5: 352, figs. 32-37. | ||||||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is based the revision of the present species by Bas (1969) and on original research of R. E. Tulloss. Bas (1969): Basidiome medium, rarely large, proportionately thickset. | ||||||||||||||||||||||||||||
| pileus | Bas (1969): 50 - 90(-130) mm wide, whitish to pale brownish, convex, dry; context white,sometimes slightly yellowish to pinkish-brownish near surface, fleshy; margin non-sulcate, appendiculate; universal veil over disc as adnate to detersile pale brown to dark brown warts ("small to rather coarse, felted-subfibrillose, subpyramidal to flat"), "towards margin gradually passing into appressed fibrillose scales." | ||||||||||||||||||||||||||||
| lamellae | Bas (1969): adnexed to free, moderately crowded to crowded, white, becoming cream to pale yellowish, sometimes with slight greenish tinge, turning reddish-brownish when bruised, with edges paler and slightly pruinose to granular and entire; lamellulae subtruncate to rounded or attenuate. | ||||||||||||||||||||||||||||
| stipe | Bas (1969): 40 - 80 × 10 - 20 mm, white, turning brownish when bruised, cylindric, mostly with attenuated base, with mostly circular zones of brownish to brown fibrillose scales; bulb sometimes with fusiform base; context solid, white,sometimes slightly yellowish to pinkish-brownish near surface; partial veil median to superior (at about one-half to three-quarters height), white, rather narrow, patent to pendent, not or faintly striate above, with edge not or slightly thickened; universal veil ?. | ||||||||||||||||||||||||||||
| odor/taste | Bas (1969): Odor indistinct or faintly pleasant, becoming slightly unpleasant. Taste mild. | ||||||||||||||||||||||||||||
| macrochemical tests |
none recorded. | ||||||||||||||||||||||||||||
| pileipellis | Bas (1969): "not well differentiated, merely an ill-delimited, dense layer between volva and trama of cap; not gelatinized." | ||||||||||||||||||||||||||||
| pileus context | not described. | ||||||||||||||||||||||||||||
| lamella trama | Bas (1969): bilateral; "with rather broad hymenopodium." | ||||||||||||||||||||||||||||
| subhymenium | Bas (1969): ramose to ?subcellular. | ||||||||||||||||||||||||||||
| basidia | Bas (1969): 40 - 70 × 9 - 13.5 (-14.5) μm, 4-sterigmate; clamps abundant. [Note: Bas noted a few basidia that he called "aberrant" and "cystidiod."—ed.] | ||||||||||||||||||||||||||||
| universal veil | Bas (1969): On pileus: elements colorless to pale yellow in alkaline solution, mostly with periclinal or interwoven orientation, over disc possibly somewhat ascending; filamentous hyphae 2 - 12 μm wide, branching; inflated cells elongate, mostly subcylindric or elongate-fusiform or subclavate, in terminal chains, 30 - 160 × 10 - 46 (-60) μm. | ||||||||||||||||||||||||||||
| stipe context | Bas (1969): longitudinally acrophysalidic; filamentous hyphae 4 - 12 μm wide, abundant, branching; acrophysalides rather abundant, mostly clavate, 80 - 200 (-300) × 18 - 45 μm; vascular hyphae scattered. | ||||||||||||||||||||||||||||
| partial veil | not described. | ||||||||||||||||||||||||||||
| lamella edge tissue | Bas (1969): "present only in very young stages"; filamentous hyphae 3 - 8 μm wide, running along (periclinal with) gill edge; inflated cells not observed. | ||||||||||||||||||||||||||||
| basidiospores |
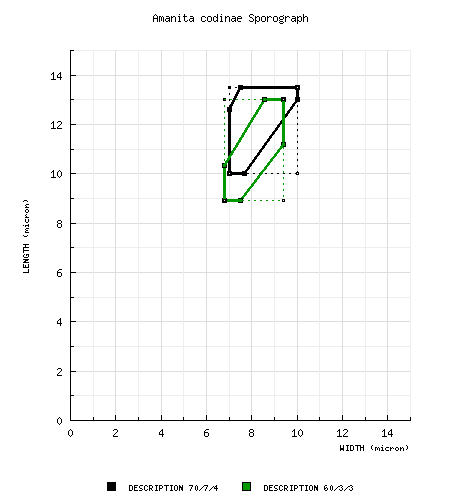
Bas (1969): [70/7/4] 10.0 - 13.5 (-14.5) × 7.0 - 10.0 (-11.0) μm, (Q = 1.30 - 1.80; Q = 1.45 - 1.60), thin-walled, amyloid, ellipsoid to elongate; apiculus not described; contents usually as large oil drop; white in deposit (becoming cream or ?"dingy yellowish." [Note: White spore deposits of amanitas almost always become cream with the passage of time.—ed.] composite from all material revised by RET: [60/3/3] (7.6-) 8.9 - 13.0 (-16.5) × (6.4-) 6.8 - 9.4 (-11.0) μm, (L = 9.7 - 11.7 μm; L' = 10.8 μm; W = 7.6 - 8.2 μm; W' = 7.9 μm; Q = (1.09-) 1.19 - 1.52 (-1.90); Q = 1.29 - 1.43; Q' = 1.36), hyaline, colorless, smooth, thin-walled, amyloid, subglobose to broadly ellipsoid to ellipsoid, infrequently elongate, often expanded at one end, usually at least somewhat adaxially flattened; apiculus sublateral, cylindric, proportionately small; contents ??; white in deposit. | ||||||||||||||||||||||||||||
| ecology |
Bas (1969): Terrestrial in fields and woods. RET: France: In grassy area by river or in mowed lawn of park or in grass of pasture without nearby trees. Italy: In meadow. | ||||||||||||||||||||||||||||
| material examined |
Bas (1969): MOROCCO: Azaghar, W of Khenifra, 8.xi.1955 Pourquier s.n. [G. Malençon 2907] (in herb. G. Malençon => MPU), notes and drawings only); Cherf el Akab, 21.xi.1964 G. Malençon 5618 (in herb. G. Malençon (MPU)); N of Rabat, littoral, 10.xi.1968 M. Guiet s.n. [G. Malençon 7002] (in herb. G. Malençon => MPU); L); Témara, 22.x.1953 G. Malençon 2550 (in herb. G. Malençon => MPU), notes and drawings only).
SPAIN: CATALUÑA—Franciac de la Selva, 22.x.1931 E. Gros s.n. (holotype, in herb. R. Maire => MPU). RET/CRC: FRANCE: MAINE-ET-LOIRE—Angers, Avrillé, "Bois du Roy," 23.ix.1995 Compres & Mme. Tricot s.n. (in herb. Jean Mornand 96 16 B1; RET 277-4); Feneu, 4.xi.1990 Marcel Galland s.n. (in herb. Jean Mornand 90 48 B1-A, as "Floccularia subcaligata"; RET 275-10); Le-Plessis-Macé, 13.xi.1990 Marcel Galland s.n. (in herb. J. Mornand 90 48 B1-B, as "Floccularia subcaligata"; RET 275-9); Sceaux d' Anjou, Grez-Neuville, 20.xi.1990 Mme. Tollafied s.n. (in herb. J. Mornand, Angers, as "Floccularia subcaligata" 90 48 B1-C; RET 275-8). ITALY: LIVORNO—Rosignano, x.1987 Ilario Filippi 1110 (in herb. I. Filippi; RET 125-7). SPAIN: MADRID—Colmenar Viejo, 22.x.2005 Fernando Cuesta s.n. (PRM; RET 498-8); Madrid, university campus, 11.x.2007 J. C. Zamora s.n. (PRM; RET 498-4, nrITS seq'd.). | ||||||||||||||||||||||||||||
| discussion |
Bas (1969): "In some of the specimens studied blackish brown matter was found in and on spores, basidia and elements of the volva; this is perhaps a result of poisoning the material with HGCl2. "Most of the microscopical characters in the description above were observed in Malençon 5618 and 7002. The type is in poor condition; only its spores and universal veil could be studied and the presences of clamps ascertained." "Amanita codinae seems to differ from A. vittadinii mainly in the brown universal veil and the usually smaller and more thickset fruiting body. Moreover it has a more strictly southern distribution. "In the few collections of A. codinae which I studied the spores are on an average somewhat more slender (average l/b 1.45 - 1.6) than those of A. vittadinii (average l/b 1.2 - 1.4). Some other differences such as the rather narrow ring and the smaller clavate cells [acrophysalides—ed.] in the tissue of the stem of A. codinae may be connected with the smaller size of the fruit-body. "Bertault (1955: 30) also mentioned a difference in the size of the spores, but that does not hold good. I found the spores of typical A. vittadinii considerably larger than he did [see "discussion" data field here—ed.]. "I have maintained Amanita codinae as a taxon on specific level also on account of Bertault's and Malençon's findings as reported by Bertault (1955: 27-32). These mycologists found A. codinae and A. vittadinii growing together abundantly, without intermediate forms. This was also reported by the author of the species, R. Maire." The following figure provides a sporograph comparison of A. codinae and A. vittadinii: | ||||||||||||||||||||||||||||
| citations | —R. E. Tulloss and C. Rodríguez Caycedo | ||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita codinae |
| name status | nomen acceptum |
| author | (R. Maire) Singer |
| english name | "Codina's Lepidella" |
| images |
1. Amanita codinae, as illustrated by C. Bas (1969). |
| drawing |
Drawing: Dr. Cornelis Bas (1969) (reproduced by courtesy of Persoonia, Leiden, the Netherlands |
| name | Amanita codinae |
| bottom links |
[ Section Lepidella page. ] [ Amanita Studies home. ] [ Keys & Checklists ] |
| name | Amanita codinae |
| bottom links |
[ Section Lepidella page. ] [ Amanita Studies home. ] [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.